Myod
| Myod1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Myod1 , myf3, myod, pum, bhlhc1, миогенная дифференциация 1, миодриф | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 159970 ; MGI : 97275 ; Гомологен : 7857 ; GeneCards : myod1 ; OMA : MYOD1 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





MYOD , также известный как определение миобластов белок 1 , [ 5 ] является белком у животных, который играет важную роль в регуляции мышечной дифференцировки . MYOD, который был обнаружен в лаборатории Гарольда М. Вайнтрауба , [ 6 ] принадлежит семейству белков, известных как миогенные регуляторные факторы (MRFS). [ 7 ] Эти BHLH (базовая спираль спирали) транскрипционные факторы действуют последовательно в миогенной дифференцировке. Члены семьи MRF MRF включают MYOD1, MYF5 , миогенон и MRF4 (MYF6). У животных, не относящихся к беспозвоночным , обычно встречается один белок MYOD.
MYOD - один из самых ранних маркеров миогенных обязательств. MYOD экспрессируется на чрезвычайно низких и по существу неопределяемых уровнях в покоящихся спутниковых клетках , но экспрессия MYOD активируется в ответ на повреждение физических упражнений или мышечной ткани. Влияние MyOD на спутниковые клетки зависит от дозы; Высокая экспрессия MYOD подавляет обновление клеток, способствует терминальной дифференцировке и может вызвать апоптоз. Хотя MYOD отмечает приверженность миобластам, развитие мышц не значительно удалено у мутантов мыши, не имеющих гена MYOD. Вероятно, это связано с функциональной избыточностью от MYF5 и/или MRF4. Тем не менее, комбинация MYOD и MYF5 жизненно важна для успеха миогенеза . [ 8 ] [ 9 ]
История
[ редактировать ]MYOD был клонирован функциональным анализом для формирования мышц, зарегистрированного в клетках в 1987 году Дэвисом, Вайнтраубом и Лассаром. Впервые он был описан как ядерный фосфопротеин в 1988 году Тапскоттом, Дэвисом, Тейером, Ченгом, Вайнтраубом и Лассаром в науке . Исследователи экспрессировали комплементарную ДНК (кДНК) белка мышиного MYOD в разных клеточных линиях ( фибробласт и адипобласт ) и обнаружили, что MYOD преобразовал их в миогенные клетки. [ 6 ] [ 10 ] В следующем году одна и та же исследовательская группа выполнила несколько тестов, чтобы определить как структуру, так и функцию белка, подтверждая их первоначальное предложение о том, что активный сайт белка состоял из спирали спиральной петли (теперь называемой базовой спиралью спиральной спирали ) для Димеризация и базовый сайт выше этой области BHLH облегчали связывание ДНК только после того, как он стал димером белка . [ 11 ] С тех пор MYOD был активной областью исследований, так как все еще относительно мало известно о многих аспектах ее функции.
Функция
[ редактировать ]Функция MYOD в разработке состоит в том, чтобы совершить мезодерму клеткам в скелетную миобласт, а затем регулировать это продолжительное состояние. MYOD также может регулировать восстановление мышц. MYOD мРНК Также сообщается, что уровни повышаются в стареющих скелетных мышцах.
Одним из основных действий MYOD является удаление клеток из клеточного цикла ( пролиферация HALT для терминальной остановки клеточного цикла в дифференцированных миоцитах) путем усиления транскрипции p21 и миогена . MYOD ингибируется циклин -зависимыми киназами ( CDK ). CDK, в свою очередь, ингибируются P21. Таким образом, MYOD повышает свою активность в клетке в виде питания.
Устойчивая экспрессия MYOD необходима для сохранения экспрессии генов, связанных с мышцами. [ 12 ]
MYOD также является важным эффектором для быстроотестовательного фенотипа мышечного волокна (типы IIA, IIX и IIB). [ 13 ] [ 14 ]
Механизмы
[ редактировать ]MYOD является фактором транскрипции и может также направлять ремоделирование хроматина посредством связывания с мотивом ДНК, известного как E-Box . Известно, что MyoD имеет связывание взаимодействий с сотнями промоторов мышечных генов и обеспечивает пролиферацию миобластов . Несмотря на то, что он не полностью понят, теперь считается, что MYOD функционирует как основной контроллер миогенеза в ассоциации включения/выключения, опосредованной FAP1 (KRAB [Krüppel-подобным Associated Box], связанным с белком 1) фосфорилированием . [ 15 ] KAP1 локализован в мышечных генах в миобластах вместе с MYOD и MEF2 (фактор усилителя транскрипции миоцитов). Здесь он служит каркасом и рекрутирует коактиваторы P300 и LSD1 , в дополнение к нескольким корепрессорам, которые включают G9A и гистон -деацетилазу HDAC1. Следствием этого набора коактиватора/корепрессора является замолчать, способствующие областям мышечных генов. Когда киназа MSK1 фосфорилирует KAP1, Corpressors, ранее связанные с каркасом, выпускаются, позволяя MYOD и MEF2 активировать транскрипцию. [ 16 ]
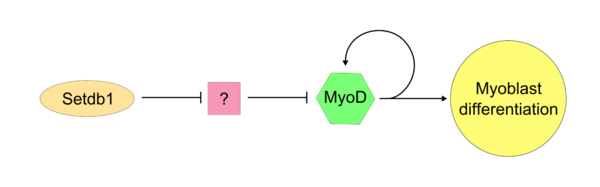
Как только MyoD «Master Controller» стал активным, SetDB1 требуется для поддержания экспрессии MYOD в ячейке. SETDB1, по -видимому, необходим для поддержания как экспрессии MYOD, так и генов, которые специфичны для мышечных тканей, поскольку снижение экспрессии SetDB1 приводит к серьезной задержке дифференцировки и определения миобластов. [ 17 ] В SETDB1 -истощенных миобластах, которые обрабатываются экзогенным MYOD, миобластическая дифференциация успешно восстанавливается. В одной модели действия SETDB1 на MYOD SETDB1 подавляет ингибитор MYOD. Этот неопознанный ингибитор, вероятно, действует конкурентно против MYOD во время типичной клеточной пролиферации. Доказательства этой модели заключаются в том, что снижение SetDB1 приводит к прямому ингибированию дифференцировки миобластов, которое может быть вызвано высвобождением неизвестного ингибитора MYOD.
Также было показано, что MYOD функционирует совместно с геном супрессора опухоли , ретинобластомой (PRB), чтобы вызвать остановку клеточного цикла в терминально дифференцированных миобластах. [ 18 ] Это делается посредством регуляции циклина , циклина D1 . Остановка клеточного цикла (в котором миобласты указывают на вывод миогенеза) зависит от непрерывной и стабильной репрессии циклина D1. Как MYOD, так и PRB необходимы для репрессии Cyclin D1, но вместо того, чтобы действовать непосредственно на циклин D1, они действуют на FRA-1, который сразу же ранний циклин D1. MYOD и PRB необходимы для подавления FRA-1 (и, следовательно, циклина D1), поскольку MYOD или PRB сами по себе недостаточно, чтобы вызвать репрессию циклина D1 и, следовательно, остановку клеточного цикла. В интронном энхансере FRA-1 были обнаружены два консервативных сайта связывания MYOD. Существует совместное действие MyoD и PRB на интронном энхансере FRA-1, который подавляет энхансер, поэтому подавляет циклин D1 и в конечном итоге приводит к остановке клеточного цикла для терминально дифференцированных миобластов. [ 19 ]
Передача сигналов Wnt может повлиять на MYOD
[ редактировать ]Было показано, что передача сигналов Wnt из соседних тканей индуцирует клетки в сомитах, которые получают эти сигналы Wnt для экспрессии PAX3 и PAX7 в дополнение к миогенным регуляторным факторам , включая MYF5 и MYOD. В частности, WNT3A может напрямую индуцировать экспрессию MYOD посредством цис-элементарных взаимодействий с дистальным энхансером и элементом ответа Wnt . [ 20 ] Wnt1 из дорсальной нейронной трубки и Wnt6/ Wnt7a из поверхностной эктодермы также участвуют в стимулировании миогенеза в сомите; Последние сигналы могут действовать в первую очередь через MYOD.
В типичных мышцах взрослых в состоянии покоя (отсутствие физиологического стресса) специфические белки семейства Wnt, которые экспрессируются, являются Wnt5a , Wnt5b, Wnt7a и Wnt4 . Когда мышца становится поврежденной (таким образом, требует регенерации) Wnt5a, Wnt5b и Wnt7a повышаются в экспрессии. По мере того, как мышца завершает восстановление Wnt7b и Wnt3a также увеличивается. Этот паттерн экспрессии Wnt в репарации мышечных клеток вызывает дифференцировку клеток -предшественников, что уменьшает количество доступных спутниковых клеток. Wnt играет решающую роль в регуляции спутниковых клеток и старении скелетных мышц, а также в регенерации. Известно, что WNT активно выражает выражение MyF5 и MYOD от Wnt1 и Wnt7a. Wnt4, Wnt5 и Wnt6 функционируют, чтобы увеличить экспрессию обоих регуляторных факторов, но на более тонком уровне. Кроме того, MYOD увеличивает WNT3A, когда миобласты подвергаются дифференциации. Активируется ли MYOD WNT посредством прямого нацеливания цис-регуляции или посредством косвенных физиологических путей, еще предстоит выяснить. [ 21 ]
Коактиваторы и подавления
[ редактировать ]IFRD1 является положительным кофактором MYOD, так как он сотрудничает с MYOD при индукции транскрипционной активности MEF2C (путем вытеснения HDAC4 из MEF2C); Кроме того, IFRD1 также подавляет транскрипционную активность NF-κB , которая, как известно, ингибирует накопление мРНК MYOD. [ 22 ] [ 23 ]
NFATC1 -это фактор транскрипции, который регулирует состав типа волокна, и переход с быстрым намотком, возникающий в результате аэробных упражнений, требует экспрессии NFATC1. Экспрессия MYOD является ключевым фактором транскрипции в волокнах быстрого подергивания, который ингибируется NFATC1 в типах окислительных волокон. NFATC1 работает, чтобы ингибировать MYOD посредством физического взаимодействия с доменом N-концевой активации MYOD, что приводит к ингибированному рекрутированию необходимого транскрипционного коактиватора P300 . NFATC1 физически нарушает взаимодействие между MYOD и P300. Это устанавливает молекулярный механизм, с помощью которого типы волокон переходят in vivo через физические упражнения с противоположными ролями для NFATC1 и MYOD. NFATC1 контролирует этот баланс путем физического ингибирования MYOD в медленных типах мышечных волокон. [ 24 ]
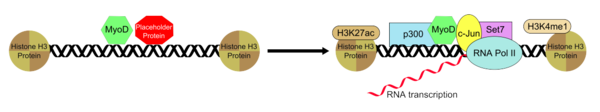
Гистондеацетилтрансфераза P300 функционирует с MYOD во взаимодействии, которое необходимо для генерации миотуба из фибробластов, которое опосредовано MYOD. Набор P300-это процесс ограничения скорости при преобразовании фибробластов в миотрубки. [ 25 ] В дополнение к P300, также известно, что MyoD рекрутирует SET7, H3K4ME1 , H3K27AC и RNAP II в энхансер, с которым связан, и это позволяет активировать мышечный ген, который является специфичным для состояния и установленным привлечением миода. Эндогенный P300, однако, необходим для функционирования MYOD, выступая в качестве важного коактиватора. MYOD ассоциативно связывается с областью усилителя в сочетании с заполнительным «предполагаемым пионерским фактором», который помогает установить и поддерживать их обоих в определенной и неактивной конформации. После удаления или инактивации на белка-заполнителе, связанном с энхансером, разрешено рекрутирование дополнительной группы транскрипционных факторов, которые помогают положительно регулировать активность энхансера, и это приводит к тому, что комплекс энхэнкер-транскрипции MYOD для принятия транскрипционного активального состояния Полем
Взаимодействия
[ редактировать ]Было показано, что MYOD взаимодействует :
- C-Jun , [ 26 ]
- CREB-связывающий белок , [ 27 ] [ 28 ]
- CSRP3 , [ 29 ]
- Циклин-зависимая киназа 4 , [ 30 ] [ 31 ]
- Циклин-зависимый ингибитор киназы 1C , [ 32 ]
- EP300 , [ 28 ] [ 33 ]
- HDAC1 , [ 34 ] [ 35 ]
- Id1 , [ 36 ] [ 37 ] [ 38 ] [ 39 ] [ 40 ] [ 41 ]
- Id2 , [ 37 ]
- Mdfi , [ 42 ]
- Не , [ 43 ]
- Белок ретинобластомы , [ 35 ] [ 44 ]
- Рецептор retinoid x альфа [ 45 ]
- STAT3 , [ 46 ] и
- TCF3 . [ 37 ] [ 47 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000129152 - Ensembl , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000009471 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «P15172 (myod1_human)» . Uniprotkb . Получено 17 июля 2019 года .
- ^ Jump up to: а беременный Дэвис Р.Л., Вайнтрауб Х, Лассар А.Б. (декабрь 1987). «Экспрессия одной трансфицированной кДНК преобразует фибробласты в миобласты». Клетка . 51 (6): 987–1000. doi : 10.1016/0092-8674 (87) 90585-x . PMID 3690668 . S2CID 37741454 .
- ^ «Ген Entrez: миогенная дифференциация MYOD1 1» .
- ^ Рудницки М.А., Шнегельсберг П.Н., Стед Р.Х., Браун Т., Арнольд Х.Х., Жениш Р. (декабрь 1993). «MYOD или MYF-5 необходим для формирования скелетных мышц». Клетка . 75 (7): 1351–1359. doi : 10.1016/0092-8674 (93) 90621-V . PMID 8269513 . S2CID 27322641 .
- ^ Hinits Y, Williams VC, Sweetman D, Donn TM, MA TP, Moens CB , et al. (Октябрь 2011). «Дефектное развитие скелета черепа, летальность личинок и гаплоинсуфона у мутантных рыбок миодов» . Девчонка Биол . 358 (1): 102–112. doi : 10.1016/j.ydbio.2011.07.015 . PMC 3360969 . PMID 21798255 .
- ^ Tapscott SJ, Davis RL, Thayer MJ, Cheng PF, Weintraub H, Lassar AB (октябрь 1988). «MYOD1: ядерный фосфопротеин, требующий гомологической области MYC для преобразования фибробластов в миобласты». Наука . 242 (4877): 405–511. Bibcode : 1988sci ... 242..405t . doi : 10.1126/science.3175662 . PMID 3175662 .
- ^ Дэвис Р.Л., Ченг П.Ф., Лассар А.Б., Тайер М., Тапскотт С., Вайнтрауб Х. (1989). «Myod и Achaete-Scute: 4-5 аминокислоты отличают миогенез от нейрогенеза». Принцесса Такамацу Симпозии . 20 : 267–278. PMID 2562185 .
- ^ Fong AP, Tapscott SJ (октябрь 2013 г.). «Программирование и перепрограммирование скелетных мышц» . Текущее мнение в области генетики и развития . 23 (5): 568–573. doi : 10.1016/j.gde.2013.05.002 . PMC 3775946 . PMID 23756045 .
- ^ Хьюз С.М., Коиши К., Рудницки М., Маггс А.М. (январь 1997). «Белок MYOD дифференциально накапливается в быстрых и медленных скелетных мышечных волокнах и требуется для нормального баланса типа волокна у грызунов» . Мех Дев . 61 (1–2): 151–163. doi : 10.1016/s0925-4773 (96) 00631-4 . PMID 9076685 . S2CID 17769090 .
- ^ Ehlers ML, Celona B, Black BL (сентябрь 2014). «NFATC1 контролирует тип скелетных мышц волокна и является отрицательным регулятором активности MYOD» . Сотовые отчеты . 8 (6): 1639–1648. doi : 10.1016/j.celrep.2014.08.035 . PMC 4180018 . PMID 25242327 .
- ^ Сингх К., Кассано М., Планета Е., Себастьян С., Джанг С.М., Сохи Г. и др. (Март 2015). «Переключатель фосфорилирования KAP1 контролирует функцию MYOD во время дифференцировки скелетных мышц» . Гены и развитие . 29 (5): 513–525. doi : 10.1101/gad.254532.114 . PMC 4358404 . PMID 25737281 .
- ^ Бакингем М., Ригби П.В. (февраль 2014 г.). «Генные регуляторные сети и транскрипционные механизмы, которые контролируют миогенез» . Ячейка развития . 28 (3): 225–238. doi : 10.1016/j.devcel.2013.12.020 . PMID 24525185 .
- ^ Song YJ, Choi JH, Lee H (февраль 2015 г.). «SETDB1 необходим для миогенной дифференцировки клеток миобластов C2C12 посредством поддержания экспрессии MYOD» . Молекулы и клетки . 38 (4): 362–372. doi : 10.14348/molcells.2015.2291 . PMC 4400312 . PMID 25715926 .
- ^ Раджаби Х.Н., Такахаши С., Эвен Мей (август 2014 г.). «Белок ретинобластомы и MYOD вместе функционируют, чтобы повторно репрессию FRA-1 и, в свою очередь, циклин D1 во время терминальной остановки клеточного цикла, связанного с миогенезом» . Журнал биологической химии . 289 (34): 23417–23427. doi : 10.1074/jbc.m113.532572 . PMC 4156083 . PMID 25006242 .
- ^ Milewska M, Grabiec K, Grzelkowska-Kowalczyk K (май 2014). «[Взаимодействие пролиферации и дифференциальных сигнальных путей в миогенезе]» . Прогресс гигиены и экспериментальной медицины . 68 : 516–526. Doi : 10.5604/1732693.1101617 . PMID 24864103 .
- ^ Pan YC, Wang XW, Teng HF, Wu YJ, Chang HC, Chen SL (февраль 2015 г.). «Пути сигналов Wnt3a активируют экспрессию MYOD, нацеливая цис-элементы внутри и снаружи его дистального усилителя» . Отчеты о биологии . 35 (2): 1–12. doi : 10.1042/bsr20140177 . PMC 4370097 . PMID 25651906 .
- ^ Motohashi N, Asakura A (январь 2014 г.). «Мышечная спутниковая гетерогенность и самообновление» . Границы в клеточной биологии и развитии . 2 (1): 1. DOI : 10.3389/fcell.2014.00001 . PMC 4206996 . PMID 25364710 .
- ^ Микели Л., Леонарди Л., Конти Ф., Буанн П., Кану Н., Карузо М. и др. (Март 2005 г.). «PC4 коактивирует MYOD путем снятия гистон-деацетилазы 4-опосредованного ингибирования фактора энхансера миоцитов 2C» . Мол Клетка. Биол . 25 (6): 2242–59. doi : 10.1128/mcb.25.6.2242-2259.2005 . PMC 1061592 . PMID 15743821 .
- ^ Микели Л., Леонарди Л., Конти Ф., Мареска Г., Колазингари С., Маттей Е. и др. (Февраль 2011 г.). «PC4/TIS7/IFRD1 стимулирует регенерацию скелетных мышц и участвует в дифференцировке миоластов как регулятор MYOD и NF-Kappab» . Дж. Биол. Химический 286 (7): 5691–707. doi : 10.1074/jbc.m110.162842 . PMC 3037682 . PMID 21127072 .
- ^ Ehlers ML, Celona B, Black BL (сентябрь 2014). «NFATC1 контролирует тип скелетных мышц волокна и является отрицательным регулятором активности MYOD» . Сотовые отчеты . 8 (6): 1639–1648. doi : 10.1016/j.celrep.2014.08.035 . PMC 4180018 . PMID 25242327 .
- ^ Sartorelli V, Huang J, Hamamori Y, Kedes L (февраль 1997 г.). «Молекулярные механизмы миогенной коактивации с помощью p300: прямое взаимодействие с доменом активации MYOD и с коробкой MADS MEF2C» . Молекулярная и клеточная биология . 17 (2): 1010–1026. doi : 10.1128/mcb.17.2.1010 . PMC 231826 . PMID 9001254 .
- ^ Бенгалия Э., Рансон Л., Шарфман Р., Дварки В.Дж., Тапскотт С.Дж., Вайнтрауб Х. и др. (Февраль 1992 г.). «Функциональный антагонизм между белками C-Jun и MYOD: прямая физическая ассоциация». Клетка . 68 (3): 507–19. doi : 10.1016/0092-8674 (92) 90187-H . PMID 1310896 . S2CID 44966899 .
- ^ Полескайя А., Нагюбнева I, Дукет А., Бенгалия Е., Робин П., Харел-Беллан А (август 2001 г.). «Взаимодействие между ацетилированным MYOD и бромодоменом CBP и/или P300» . Мол Клетка. Биол . 21 (16): 5312–20. doi : 10.1128/mcb.21.16.5312-5320.2001 . PMC 87255 . PMID 11463815 .
- ^ Jump up to: а беременный Sartorelli V, Huang J, Hamamori Y, Kedes L (февраль 1997 г.). «Молекулярные механизмы миогенной коактивации с помощью p300: прямое взаимодействие с доменом активации MYOD и с коробкой MADS MEF2C» . Мол Клетка. Биол . 17 (2): 1010–26. doi : 10.1128/mcb.17.2.1010 . PMC 231826 . PMID 9001254 .
- ^ Kong Y, Flick MJ, Kudla AJ, Konieczny SF (август 1997). «Белок мышц LIM способствует миогенезу, усиливая активность MYOD» . Мол Клетка. Биол . 17 (8): 4750–60. doi : 10.1128/mcb.17.8.4750 . PMC 232327 . PMID 9234731 .
- ^ Zhang JM, Zhao X, Wei Q, Paterson BM (декабрь 1999 г.). «Прямое ингибирование активности CDK -киназы G (1) MYOD способствует отмене клеточного цикла миобластов и дифференцировки терминала» . Embo j . 18 (24): 6983–93. doi : 10.1093/emboj/18.24.6983 . PMC 1171761 . PMID 10601020 .
- ^ Zhang JM, Wei Q, Zhao X, Paterson BM (февраль 1999 г.). «Соединение клеточного цикла и миогенез через циклин D1-зависимое взаимодействие MyOD с CDK4» . Embo j . 18 (4): 926–33. doi : 10.1093/emboj/18.4.926 . PMC 1171185 . PMID 10022835 .
- ^ Reynaud EG, Leibovitch MP, Tintignac LA, Pelpel K, Guillier M, Leibovitch SA (июнь 2000 г.). «Стабилизация MYOD путем прямого связывания с p57 (KIP2)» . Дж. Биол. Химический 275 (25): 18767–76. doi : 10.1074/jbc.m907412199 . PMID 10764802 .
- ^ Лау П., Бейли П., Доухан Д.Х., Маскат Г.Е. (январь 1999 г.). «Экзогенная экспрессия доминантного отрицательного вектора Roralpha1 в мышечных клетках нарушает дифференцировку: Roralpha1 напрямую взаимодействует с p300 и myod» . Нуклеиновые кислоты Res . 27 (2): 411–20. doi : 10.1093/nar/27.2.411 . PMC 148194 . PMID 9862959 .
- ^ Puri PL, Iezzi S, Stiegler P, Chen TT, Schiltz RL, Muscat GE и др. (Октябрь 2001 г.). «Гистоновые деацетилазы класса I последовательно взаимодействуют с MYOD и PRB во время скелетного миогенеза» . Мол Клетка . 8 (4): 885–97. doi : 10.1016/s1097-2765 (01) 00373-2 . PMID 11684023 .
- ^ Jump up to: а беременный Mal A, Sturniolo M, Schiltz RL, Ghosh Mk, Harter ML (апрель 2001 г.). «Роль гистондеацетилазы HDAC1 в модуляции транскрипционной активности MYOD: ингибирование миогенной программы» . Embo j . 20 (7): 1739–53. doi : 10.1093/emboj/20.7.1739 . PMC 145490 . PMID 11285237 .
- ^ Garkavtsev I, Kozin SV, Chernova O, Xu L, Winkler F, Brown E, et al. (Март 2004 г.). «Белок -супрессор -супрессор кандидата 4 регулирует рост опухоли головного мозга и ангиогенез». Природа . 428 (6980): 328–32. Bibcode : 2004natur.428..328G . doi : 10.1038/nature02329 . PMID 15029197 . S2CID 4427531 .
- ^ Jump up to: а беременный в Langlands K, Yin X, Anand G, Prochownik EV (август 1997 г.). «Дифференциальные взаимодействия белков ID с факторами транскрипции с базовой спиралью» . Дж. Биол. Химический 272 (32): 19785–93. doi : 10.1074/jbc.272.32.19785 . PMID 9242638 .
- ^ Finkel T, Duc J, Fearon ER, Dang CV, Tomaselli GF (январь 1993 г.). «Обнаружение и модуляция in vivo взаимодействия белка-белка-спираль-спираль» » . Дж. Биол. Химический 268 (1): 5–8. doi : 10.1016/s0021-9258 (18) 54105-3 . PMID 8380166 .
- ^ Гупта К., Ананд Г., Инь Х, Гроув Л., Прохоун Э.В. (март 1998 г.). «MMIP1: новый белок лейциновой молнии, который изменяет подавляющее воздействие членов сумасшедшего семейства на c-myc» . Онкоген . 16 (9): 1149–59. doi : 10.1038/sj.onc.1201634 . PMID 9528857 .
- ^ McLoughlin P, Ehler E, Carlile G, Licht JD, Schäfer BW (октябрь 2002 г.). «Белковой Dral/FHL2, только Limly, взаимодействует с и является корепрессором для белка цинкового пальца с промилоцитарным лейкозом» . Дж. Биол. Химический 277 (40): 37045–53. doi : 10.1074/jbc.m203336200 . PMID 12145280 .
- ^ Ling MT, Chiu YT, Lee TK, Leung SC, Fung MK, Wang X, et al. (Сентябрь 2008 г.). «ID-1 индуцирует протеасом-зависимую деградацию белка HBX». J. Mol. Биол . 382 (1): 34–43. doi : 10.1016/j.jmb.2007.06.020 . PMID 18674781 .
- ^ Chen CM, Kraut N, Groudine M, Weintraub H (сентябрь 1996 г.). «I-MF, новый миогенный репрессор, взаимодействует с членами семьи MYOD» . Клетка . 86 (5): 731–41. doi : 10.1016/s0092-8674 (00) 80148-8 . PMID 8797820 . S2CID 16252710 .
- ^ Ленорманд Дж. Л., Бенаюн Б., Гилье М., Вандромм М., Лейбович М.П., Лейбович С.А. (февраль 1997 г.). «MOS активирует миогенную дифференцировку, способствуя гетеродимеризации белков MYOD и E12» . Мол Клетка. Биол . 17 (2): 584–93. doi : 10.1128/mcb.17.2.584 . PMC 231783 . PMID 9001211 .
- ^ Gu W, Schneider JW, Condorelli G, Kaushal S, Mahdavi V, Nadal-Ganard B (февраль 1993 г.). «Взаимодействие миогенных факторов и белка ретинобластомы опосредует приверженность и дифференцировку мышечных клеток». Клетка . 72 (3): 309–24. doi : 10.1016/0092-8674 (93) 90110-c . PMID 8381715 . S2CID 21581966 .
- ^ Froschlé A, Alric S, Kitzmann M, Carnac G, Auradé F, Rochette-Egly C, et al. (Июль 1998 г.). «Рецепторы ретиноевой кислоты и мышечные белки B-HLH: партнеры в миогенезе, вызванном ретиноидом» . Онкоген . 16 (26): 3369–78. doi : 10.1038/sj.onc.1201894 . PMID 9692544 .
- ^ Kataoka Y, Matsumura I, Ezoe S, Nakata S, Takigawa E, Sato Y, et al. (Ноябрь 2003). «Взаимное торможение между MYOD и STAT3 в регуляции роста и дифференцировки миобластов» . Дж. Биол. Химический 278 (45): 44178–87. doi : 10.1074/jbc.m304884200 . PMID 12947115 .
- ^ Малеки С.Дж., Ройер К.А., Херлбурт Б.К. (июнь 1997 г.). «Гетеродимеры MYOD-E12 и гомодимеры MYOD-Myod одинаково стабильны». Биохимия . 36 (22): 6762–7. doi : 10.1021/bi970262m . PMID 9184158 .
Внешние ссылки
[ редактировать ]- MYOD+белок в Национальной библиотеке Медицинской библиотеки Медицинской библиотеки (Mesh)
- Обзор всей структурной информации, доступной в PDB для Uniprot : P10085 (белок 1 мыши миобластов) в PDBE-KB .
PDB Галерея |
|---|
