Ядерный рецептор

В области молекулярной биологии ядерные рецепторы представляют собой класс белков, ответственных за восприятие стероидов , щитовидной железы гормонов , витаминов и некоторых других молекул. Эти внутриклеточные рецепторы работают с другими белками, чтобы регулировать экспрессию специфических генов , тем самым контролируя развитие , гомеостаз и метаболизм организма.
Ядерные рецепторы связываются непосредственно с ДНК, регулирующей экспрессию соседних генов; Следовательно, эти рецепторы классифицируются как факторы транскрипции . [ 2 ] [ 3 ] Регуляция экспрессии генов ядерными рецепторами часто происходит в присутствии лиганда - молекулы, которая влияет на поведение рецептора. Связывание лиганда с ядерным рецептором приводит к конформационному изменению, активирующим рецептор. Результатом является повышение или понижающая регуляция экспрессии генов.
Уникальным свойством ядерных рецепторов, которое отличает их от других классов рецепторов, является их прямой контроль геномной ДНК. Ядерные рецепторы играют ключевую роль как в эмбриональном развитии, так и в гомеостазе взрослых. Как обсуждалось ниже, ядерные рецепторы классифицируются в соответствии с механизмом [ 4 ] [ 5 ] или гомология . [ 6 ] [ 7 ]
Виды распределение
[ редактировать ]Ядерные рецепторы специфичны для метазоя (животных) и не обнаруживаются в протистах , водорослях , грибах или растениях. [ 8 ] Среди ранних линий животных с секвенированными геномами с секвенированными геномами два были зарегистрированы из Амфимедон Квинсландия , два из расчески губки [ 9 ] Четыре из плаконов Trichoplax Adhaerens и 17 из Cniidarian Nematostella Vectensis [ 10 ] Только 270 ядерных рецепторов только Caenorhabditis Elegans , [ 11 ] 21 в фруктовой мух и других насекомых, [ 12 ] 73 у рыбок данио . [ 13 ] Люди, мыши и крысы имеют соответственно 48, 49 и 47 ядерных рецепторов каждый. [ 14 ]
Лиганды
[ редактировать ]
Лиганды, которые связываются и активируют ядерные рецепторы, включают липофильные вещества, такие как эндогенные гормоны , витамины A и D и ксенобиотические гормоны . Поскольку экспрессия большого количества генов регулируется ядерными рецепторами, лиганды, которые активируют эти рецепторы, могут оказывать глубокое влияние на организм. Многие из этих регулируемых генов связаны с различными заболеваниями, что объясняет, почему молекулярные мишени приблизительно 13% Управления по проживанию и лекарствам США (FDA) одобрены ядерными рецепторами. [ 15 ]
Ряд ядерных рецепторов, называемых рецепторами сирот , [ 16 ] не иметь известных (или, по крайней мере, в целом согласованы) эндогенные лиганды. Некоторые из этих рецепторов, таких как FXR , LXR и PPAR, связывают ряд метаболических промежуточных соединений, таких как жирные кислоты, желчные кислоты и/или стеролы с относительно низкой аффинностью. Следовательно, эти рецепторы могут функционировать как метаболические датчики. Другие ядерные рецепторы, такие как CAR и PXR , по-видимому, функционируют как ксенобиотические датчики, повышающие экспрессию ферментов цитохрома P450 , которые метаболизируют эти ксенобиотики. [ 17 ]
Структура
[ редактировать ]Большинство ядерных рецепторов имеют молекулярные массы от 50 000 до 100 000 далтонов .
Ядерные рецепторы имеют модульные структуры и содержат следующие домены : [ 18 ] [ 19 ]
- (AB) N-концевой регуляторный домен: содержит функцию активации 1 ( AF-1 ), действие которой не зависит от наличия лиганда. [ 20 ] Транскрипционная активация AF-1, как правило, очень слаба, но она синергирует с AF-2 в электронном домене (см. Ниже), чтобы повысить более высокую активацию экспрессии генов. Домен AB сильно варьируется в последовательности между различными ядерными рецепторами.
- (C) ДНК-связывающий домен ( DBD ): высоко консервативный домен, содержащий два цинковых пальца , которые связываются с специфическими последовательностями ДНК, называемых элементами отклика гормонов (HRE). Недавно новый мотив цинкового пальца (CHC2) идентифицируется в паразитных плоских червях NRS. [ 21 ]
- (D) Перекрываемая область: считается гибким доменом, который соединяет DBD с LBD. Влияет на внутриклеточный перевозки и субклеточное распределение с последовательности пептидов -мишени .
- (E) Домен связывания лиганда ( LBD ): умеренно консервативный по последовательности и высоко консервативный по структуре между различными ядерными рецепторами. Структура . LBD называется альфа -спиральной сэндвич -складом , в которой три антипотативные альфа -спирали («сэндвич -наполнение») окружены двумя альфа -спиралями с одной стороны и три с другой («хлеб») Половая лиганда находится внутри внутренней части LBD и чуть ниже трех антипокано -параллельных спиральных спиральных сэндвич «наполнение». Наряду с DBD, LBD вносит свой вклад в границу димеризации рецептора и, кроме того, связывает коактиватор и корепрессора белки . LBD также содержит функцию активации 2 ( AF-2 ), действие которой зависит от присутствия связанного лиганда, контролируемой конформацией спирали 12 (H12). [ 20 ]
- (F) C-концевой домен: сильно варьируется в последовательности между различными ядерными рецепторами.
Домены ДНК-связывания (C) и связывания лиганда (E) являются независимо сложенными и структурно стабильными, в то время как N-концевые (A/B), петля (D) и необязательные C-концевые (F) домены могут быть конформационно Гибкий и беспорядочный. [ 22 ] Относительные ориентации доменов очень различаются, сравнивая три известные многодоменные кристаллические структуры, две из которых связывают на DR1 (DBD, разделенные на 1 п.н.), [ 1 ] [ 23 ] Одно связывание на DR4 (на 4 п.н.). [ 24 ]
Верх - схематическая аминокислотная последовательность ядерного рецептора. Внизу - 3D -структуры областей DBD (связанных с ДНК) и LBD (связанными с гормонами) областей ядерного рецептора. Показанные структуры относятся к рецептору эстрогена . Экспериментальные структуры N-концевого домена (A/B), шарнирной области (D) и C-концевой домен (F) не были определены, поэтому представлены красными, фиолетовыми и оранжевыми пунктирными линиями соответственно. |
|
| ||||||||||||||||||||||||||||||||||||||||||||||

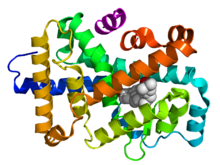
Механизм действия
[ редактировать ]Ядерные рецепторы представляют собой многофункциональные белки, которые трансдуцируют сигналы их родственных лигандов . Ядерные рецепторы (NRS) могут быть классифицированы на два широких класса в соответствии с их механизмом действия и субклеточного распределения в отсутствие лиганда.
Маленькие липофильные вещества, такие как природные гормоны, диффундируют через клеточную мембрану и связываются с ядерными рецепторами, расположенными в цитозоле (NR типа I) или ядре (NR типа II) клетки. Связывание вызывает конформационное изменение в рецепторе, которое, в зависимости от класса рецептора, запускает каскад нисходящих событий, которые направляют NRS к сайтам регуляции транскрипции ДНК, которые приводят к повышению или пониженной экспрессии генов. Они обычно функционируют как гомо/гетеродимеры. [ 27 ] Кроме того, два дополнительных класса, тип III, которые являются вариантом типа I, и типа IV, которые связывают ДНК в качестве мономеров, также были идентифицированы. [ 4 ]
Соответственно, ядерные рецепторы могут быть разделены на следующие четыре механистические классы: [ 4 ] [ 5 ]
Тип i
[ редактировать ]Связывание лиганда с ядерными рецепторами типа I в цитозоле приводит к диссоциации белков теплового шока , гомомеризации , транслокации ( IE , активного транспорта ) из цитоплазмы в ядро клеток и связывание с специфическими последовательностями ДНК, известных как элементы ответа гормона (HRES). Ядерные рецепторы типа I связываются с HRE, состоящими из двух половинов, разделенных переменной длиной ДНК, а вторая половина сайта имеет последовательность, перевернутую от первого (инвертированный повтор). Ядерные рецепторы типа I включают членов подсемейства 3, таких как рецептор андрогена , рецепторы эстрогена , рецептор глюкокортикоидов и рецептор прогестерона . [ 28 ]
Было отмечено, что некоторые из ядерных рецепторов NR подсем 2 могут связываться с прямым повторением вместо инвертированного повторного HRE. Кроме того, некоторые ядерные рецепторы, которые связывают либо как мономеры, так и димеры, только с одним доменом связывания ДНК рецептора, прикрепленным к одному половине сайта HRE. Эти ядерные рецепторы считаются рецепторами сирот , поскольку их эндогенные лиганды до сих пор неизвестны.
ядерного рецептора/ДНК Затем комплекс рекрутирует другие белки, которые транскрибируют ДНК вниз по течению от HRE в РНК мессенджера и в конечном итоге белок , что вызывает изменение функции клеток.
Тип II
[ редактировать ]Рецепторы типа II, в отличие от типа I, сохраняются в ядре независимо от состояния связывания лиганда и, кроме того, связываются как гетеродимерные (обычно с RXR ) с ДНК. [ 27 ] В отсутствие лиганда ядерные рецепторы типа II часто комплексны с корепрессора белками . Связывание лиганда с ядерным рецептором вызывает диссоциацию корепрессора и рекрутирование белков коактиватора . Дополнительные белки, включая РНК -полимеразу, затем рекрутируются в комплекс NR/ДНК, который транскрибирует ДНК в РНК мессенджера.
Ядерные рецепторы типа II включают в основном подсемейство 1, например, рецептор ретиноевой кислоты , рецептор ретиноида X и рецептор гормонов щитовидной железы . [ 29 ]
Тип III
[ редактировать ]Ядерные рецепторы типа III (в основном NR подсемейство 2) аналогичны рецепторам I типа I в том, что оба класса связываются с ДНК в качестве гомодимеров. Однако ядерные рецепторы типа III, в отличие от типа I, связываются с прямым повторением вместо инвертированного повторного HRE.
Тип IV
[ редактировать ]Ядерные рецепторы типа IV связываются в виде мономеров или димеров, но только один домен связывающего ДНК рецептора связывается с HRE с одной половиной. Примеры рецепторов типа IV обнаружены в большинстве подсемейств NR.
Димеризация
[ редактировать ]Человеческие ядерные рецепторы способны к димеризации со многими другими ядерными рецепторами (гомотипическая димеризация), как было показано из крупномасштабных экспериментов Y2H и усилий по добыче текста литературы, которые были сосредоточены на конкретных взаимодействиях. [ 30 ] [ 31 ] [ 27 ] Тем не менее, существует специфичность, когда члены той же подсемейства имеют очень похожие партнеры по димеризации NR, а базовая димеризационная сеть имеют определенные топологические особенности, такие как наличие высоко связанных концентраторов (RXR и SHP). [ 27 ]
Coregulatory Belotins
[ редактировать ]Ядерные рецепторы, связанные с элементами реакции гормона, набирают значительное количество других белков (называемых корегуляторами транскрипции ), которые облегчают или ингибируют транскрипцию связанного гена -мишеня в мРНК. [ 32 ] [ 33 ] [ 34 ] Функция этих корегуляторов варьируется и включает в себя ремоделирование хроматина (делая ген -мишень более или менее доступным для транскрипции), либо мостичной функции для стабилизации связывания других корегуляторных белков. Ядерные рецепторы могут специфично связываться с рядом белков корегулятора и тем самым влиять на клеточные механизмы передачи сигнала как напрямую, так и косвенно. [ 35 ]
Коактиваторы
[ редактировать ]Связывание агонистских лигандов (см. Раздел ниже) с ядерными рецепторами индуцирует конформацию рецептора, которая предпочтительно связывает коактиваторные белки. Эти белки часто обладают внутренней активностью гистонцетилтрансферазы (HAT), которая ослабляет связь гистонов с ДНК и, следовательно, способствует транскрипции генов.
Corpressors
[ редактировать ]Связывание антагонистов лигандов с ядерными рецепторами в отличие от конформации рецептора, которая предпочтительно связывает белки корепрессора . Эти белки, в свою очередь, рекрутируют гистондеацетилазы (HDAC), которые укрепляют связь гистонов с ДНК и, следовательно, подавляют транскрипцию генов.
Агонизм против антагонизма
[ редактировать ]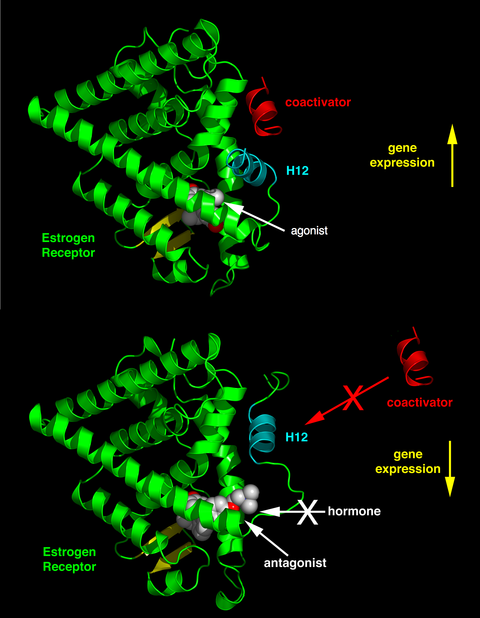
В зависимости от вовлеченного рецептора, химическая структура лиганда и затронутой ткани, лиганды ядерных рецепторов могут проявлять существенные различные эффекты, начиная с спектра от агонизма до антагонизма до обратного агонизма. [ 38 ]
Агонисты
[ редактировать ]Активность эндогенных лигандов (таких как гормоны эстрадиола и тестостерона ), когда он связан с их родственными ядерными рецепторами, обычно для повышения регулирования экспрессии генов. Эта стимуляция экспрессии генов лигандом называется агонистским ответом. Агонистические эффекты эндогенных гормонов также можно имитировать определенными синтетическими лигандами, например, глюкокортикоидного рецептора противовоспалительным лекарственным препаратом дексаметазона . Агонистские лиганды работают, вызывая конформацию рецептора, который способствует связыванию коактиватора (см. Верхую половину рисунка справа).
Антагонисты
[ редактировать ]Другие лиганды синтетических ядерных рецепторов не оказывают очевидного влияния на транскрипцию генов в отсутствие эндогенного лиганда. Однако они блокируют влияние агониста посредством конкурентного связывания с одним и тем же сайтом связывания в ядерном рецепторе. Эти лиганды называются антагонистами. Примером антагонистического лекарственного средства ядерного рецептора является мифепристон , который связывается с рецепторами глюкокортикоида и прогестерона и, следовательно, блокирует активность эндогенного гормонного кортизола и прогестерона соответственно. Антагонисты лиганды работают, индуцируя конформацию рецептора, которая предотвращает связывание коактиватора, и способствует связыванию корепрессора (см. Нижнюю половину рисунка справа).
Обратные агонисты
[ редактировать ]Наконец, некоторые ядерные рецепторы способствуют низкому уровню транскрипции генов в отсутствие агонистов (также называемых базальной или конститутивной активностью). Синтетические лиганды, которые снижают этот базальный уровень активности в ядерных рецепторах, известны как обратные агонисты . [ 39 ]
Селективные рецепторные модуляторы
[ редактировать ]Ряд лекарств, которые работают через ядерные рецепторы, демонстрируют агонистский ответ в некоторых тканях и антагонистический ответ в других тканях. Такое поведение может иметь существенные преимущества, поскольку оно может позволить сохранить желаемые полезные терапевтические эффекты препарата при минимизации нежелательных побочных эффектов. Препараты с этим смешанным агонистом/антагонистовым профилем действия называются селективными рецепторными модуляторами (SRMS). Примеры включают селективные модуляторы андрогенных рецепторов ( SARM ), селективные модуляторы рецепторов эстрогена ( SERM ) и селективные модуляторы рецепторов прогестерона ( SPRM ). Механизм действия SRM может варьироваться в зависимости от химической структуры лиганда и вовлеченного рецептора, однако считается, что многие SRM работают, способствуя конформации рецептора, который тесно сбалансирован между агонизмом и антагонизмом. В тканях, где концентрация коактиваторных белков выше, чем корепрессоры , равновесие смещается в направлении агониста. Наоборот в тканях, где Доминируют корепрессоры , лиганд ведет себя как антагонист. [ 40 ]
Альтернативные механизмы
[ редактировать ]
Трансферропрессия
[ редактировать ]Наиболее распространенный механизм действия ядерного рецептора включает прямое связывание ядерного рецептора с элементом отклика Гормона ДНК. Этот механизм называется трансактивацией . Однако некоторые ядерные рецепторы обладают не только способностью напрямую связываться с ДНК, но и с другими факторами транскрипции. Это связывание часто приводит к дезактивации второго фактора транскрипции в процессе, известном как трансрепрессия . [ 41 ] Одним из примеров ядерного рецептора, способного транспрессии, является глюкокортикоидный рецептор (GR). Кроме того, некоторые лиганды GR, известные как селективные глюкокортикоидные агонисты рецепторов ( SEGRA ), способны активировать GR таким образом, чтобы GR более сильно транспрессирует, чем трансактивирует. Эта селективность увеличивает разделение между желаемыми противовоспалительными эффектами и нежелательными метаболическими побочными эффектами этих селективных глюкокортикоидов .
Не геномный
[ редактировать ]Классическое прямое влияние ядерных рецепторов на регуляцию генов обычно занимает несколько часов до того, как функциональный эффект наблюдается в клетках из -за большого количества промежуточных стадий между активацией ядерного рецептора и изменениями в уровнях экспрессии белка. Однако было отмечено, что многие эффекты применения ядерных гормонов, такие как изменения активности ионных каналов, возникают в течение нескольких минут, что противоречит классическому механизму действия ядерного рецептора. В то время как молекулярная мишень для этих негеномных эффектов ядерных рецепторов не была окончательно продемонстрирована, было предположительно, что существуют варианты ядерных рецепторов, которые связаны с мембраной, вместо того, чтобы локализоваться в цитозоле или ядре. Кроме того, эти рецепторы, связанные с мембраной, функционируют посредством альтернативных механизмов трансдукции сигнала , не включающих регуляцию генов. [ 42 ] [ 43 ]
Хотя было предположено, что существует несколько рецепторов, связанных с мембранами для ядерных гормонов, было показано, что многие из быстрых эффектов требуют канонических ядерных рецепторов. [ 44 ] [ 45 ] Однако проверка относительной важности геномных и негеномных механизмов in vivo было предотвращено отсутствием специфических молекулярных механизмов для негеномных эффектов, которые могут быть заблокированы мутацией рецептора без нарушения его прямого воздействия на экспрессию генов.
Молекулярный механизм негеномной передачи сигналов через рецептор ядерного гормона щитовидной железы TRβ включает фосфатидилинозитол 3-киназу ( PI3K ). [ 46 ] Эта передача сигналов может быть заблокирована одним тирозином на фенилаланиновую замену в TRβ без нарушения прямой регуляции генов. [ 47 ] Когда мыши были созданы с помощью этой единственной, консервативной аминокислотной замены в TRβ, [ 47 ] Синаптическое созревание и пластичность в гиппокампе были нарушены почти так же эффективно, как полностью блокировка синтеза гормонов щитовидной железы. [ 48 ] Этот механизм, по -видимому, сохраняется у всех млекопитающих, но не у TRα или в любых других ядерных рецепторах. Таким образом, фосфотирозин-зависимая ассоциация TRβ с PI3K обеспечивает потенциальный механизм для интеграции регуляции развития и метаболизма с помощью гормонов щитовидной железы и рецепторных тирозинкиназ. Кроме того, передача сигналов гормона щитовидной железы через PI3K может изменить экспрессию генов. [ 49 ]
Члены семьи
[ редактировать ]Ниже приведен список 48 известных ядерных рецепторов человека (и их ортологи у других видов) [ 14 ] [ 50 ] [ 51 ] категоризируется в соответствии с гомологией последовательности . [ 6 ] [ 7 ] Список также включает отдельных членов семьи, у которых не хватает человеческих ортологов (символ NRNC, выделенный желтым цветом).
| Подсемейство | Группа | Член | ||||||
|---|---|---|---|---|---|---|---|---|
| NRNC символ [ 6 ] | Аббревиатура | Имя | Ген | Лиганд (ы) | ||||
| 1 | Рецептор щитовидной железы, подобный рецептору | А | Рецептор гормонов щитовидной железы | NR1A1 | TRα | Гормоны щитовидной железы рецептор-α | Через | гормон щитовидной железы |
| NR1A2 | TRβ | Рецептор гормонов щитовидной железы β | Трэнд | |||||
| Беременный | Рецептор ретиноевой кислоты | NR1B1 | Rarα | Рецептор ретиноевой кислоты-α | НИКОГДА | витамин А и связанные с ними соединения | ||
| NR1B2 | Rarβ | Рецептор ретиноевой кислоты-β | РАРБ | |||||
| NR1B3 | Рарг | Рецептор ретиноевой кислоты-γ | Рарг | |||||
| В | Пероксисом-пролифератор-активированный рецептор | NR1C1 | PPARα | Пероксисом-пролифератор-активированный рецептор-α | Ppara | жирные кислоты , простагландины | ||
| NR1C2 | PPAR-B/D. | Пероксисом-пролифератор-активированный рецептор-β/Δ | Ppard | |||||
| NR1C3 | PPARγ | Пероксисом-пролифератор-активированный рецептор-γ | PPARG | |||||
| Дюймовый | Rev-Four | NO1D1 | Rev-Erbaα | Rev-Erbaα | NO1D1 | гем | ||
| NR1D2 | Rev-Erbaβ | Rev-Erbaα | NR1D2 | |||||
| И | E78c-подобный (членистоногие, трематод, муллос, нематода) [ 50 ] [ 52 ] |
NR1E1 | EIP78C | Индуцированный экдизоном белок 78c | EIP78C | |||
| Фон | Связанный с RAR рецептор сирот | NR1F1 | RORα | Связанный с RAR-рецептор-сирот-α | Рора | холестерин , атры | ||
| NR1F2 | RORβ | Связанный с РАР-рецептор-сирот-β | Рорб | |||||
| NR1F3 | RORγ | Связанный с RAR-рецептор-сирот-γ | Орк | |||||
| Глин | CNR14-подобный (нематода) [ 50 ] | NR1G1 | Секс-1 | Рецептор стероидных гормонов CNR14 [ 53 ] | Секс-1 | |||
| ЧАС | Печень x рецептор -похожий на | WR1H1 | Экземпляр | Рецептор экдизона, ECR (членисторонний) | Экземпляр | экдистероиды | ||
| NR1H2 | LXRβ | Рецептор печени x | NR1H2 | оксистеролы | ||||
| WR1H3 | LXRα | Печень X Рецептор-α | WR1H3 | |||||
| NR1H4 | FXR | Фарнероидный X Рецептор | NR1H4 | |||||
| NR1H5 [ 54 ] | FXR-B | Фарнероидный X Рецептор-β (псевдоген у человека) |
NR1H5P | |||||
| я | Рецептор витамина D. | NR1I1 | Vdr | Рецептор витамина D. | Vdr | Витамин d | ||
| NR1I2 | Pxr | Беременная x рецептор | NR1I2 | Ксенобиотики | ||||
| NR1I3 | МАШИНА | Конститутивный рецептор андростана | NR1I3 | Андростан | ||||
| Дж | HR96-подобный [ 50 ] | NR1J1 | HR96/ DAF-12 | Рецептор ядерного гормона HR96 | HR96 | холестерин / дафахроновая кислота [ 55 ] | ||
| NR1J2 | ||||||||
| NR1J3 | ||||||||
| K | HR1-подобный [ 50 ] | NR1K1 | HR1 | Рецептор ядерного гормона HR1 | ||||
| 2 | Рецептор-рецептор-рецептор | А | Гепатоцит ядерный фактор-4 | NO2A1 | HNF4α | Гепатоцит ядерный фактор-4-α | HHF4A | жирные кислоты |
| NR2A2 | HNF4γ | Гепатоцит ядерный фактор-4-γ | HNF4G | |||||
| Беременный | Рецептор ретиноида X. | NR2B1 | RXRα | Ретиноид X-рецептор-α | Rxra | ретиноиды | ||
| NR2B2 | RXRβ | Ретиноидный X Рецептор-β | Rxrb | |||||
| NR2B3 | RXRγ | Ретиноид X-рецептор-γ | Rxrg | |||||
| NR2B4 | USP | Ультраспиральный белок (членисторонний) | USP | фосфолипиды [ 56 ] | ||||
| В | Рецептор яичка | NR2C1 | TR2 | Рецептор яичка 2 | NR2C1 | |||
| NR2C2 | TR4 | Рецептор яичка 4 | NR2C2 | |||||
| И | TLX/PNR | NR2E1 | TLX | Гомолог гена Drosophila Hailless | NR2E1 | |||
| NR2E3 | Пнр | Фоторецептор-специфический ядерный рецептор | NR2E3 | |||||
| Фон | Переворот /ухо | NR2F1 | Coup-TFI | Цыпленок овальбумин вверх по течению промотор-транскрипционный фактор I | NR2F1 | |||
| NR2F2 | Coup-tfii | Куриный овальбумин вверх по течению промотор-транскрипционный фактор II | NR2F2 | ретиноевая кислота (слабая) [ 57 ] | ||||
| NR2F6 | Ухо-2 | V-erbA-related | NR2F6 | |||||
| 3 | Рецептор эстрогена | А | Рецептор эстрогена | NR3A1 | ERα | Рецептор эстрогена α | ESR1 | эстрогены |
| NR3A2 | ERβ | Рецептор эстрогена β | ESR2 | |||||
| Беременный | Рецептор, связанный с эстрогеном | NR3B1 | ERRα | Связанный с эстрогеном рецептор-α | Эсрра | |||
| NR3B2 | Ошибка | Связанный с эстрогеном рецептор-β | ESRRB | |||||
| NR3B3 | Ошибка | Связанный с эстрогеном рецептор-γ | Esrrg | |||||
| В | 3-кетостероидные рецепторы | NR3C1 | Гр | Глюкокортикоидный рецептор | NR3C1 | кортизол | ||
| NR3C2 | МИСТЕР | Минералокортикоидный рецептор | NR3C2 | альдостерон | ||||
| NR3C3 | Премьер -министр | Рецептор прогестерона | Пг | прогестерон | ||||
| NR3C4 | С | Андрогенный рецептор | С | тестостерон | ||||
| Дюймовый | Рецептор эстрогена (в лофотрохозооа ) [ 58 ] |
NR3D | ||||||
| И | Рецептор эстрогена (в Cnidaria ) [ 59 ] |
NR3E | ||||||
| Фон | Рецептор эстрогена (в Placozoa ) [ 59 ] |
NR3F | ||||||
| 4 | Фактор роста нерва IB-подобного | А | Ngfib/nurr1/nor1 | NR4A1 | Нгфиб | Фактор роста нерва IB | NR4A1 | |
| NR4A2 | Nurr1 | Связанный с ядерным рецептором 1 | NR4A2 | |||||
| NR4A3 | Нор | Нейрон-рецептор 1-го сироты 1 | NR4A3 | |||||
| 5 | Стероидогенный Фактор, похожий на |
А | SF1/LRH1 | NR5A1 | SF1 | Стероидогенный фактор 1 | NR5A1 | фосфатидилинозитолы |
| NR5A2 | LRH-1 | Рецептор печени гомолог-1 | NR5A2 | фосфатидилинозитолы | ||||
| Беременный | HR39-подобный | NR5B1 [ 50 ] | HR39/ FTZ-F1 | Рецептор ядерного гормона Fushi Tarazu Фактор я бета | HR39 | |||
| 6 | Ядерный фактор, похожий на зародышевой клетки | А | GCNF | NR6A1 | GCNF | Ядерный фактор зародышевых клеток | NR6A1 | |
| 7 | NR с двумя доменами связывания ДНК [ 60 ] [ 50 ] [ 61 ] | А | 2DBD-NRα | NR7A1 | 2DBD-NRA2 | |||
| Беременный | 2DBD-NRβ | NR7B1 | 2DBD-NRA3 | |||||
| В | 2DBD-NRγ | NR7C1 | 2DBD-NRA1 | членистоногие "α/β" | ||||
| 8 | NR8 [ 62 ] ( Eumetao ) | А | NR8A | NR8A1 | CGNR8A1 | Ядерный рецептор 8 | AKG49571 | |
| 0 | Разное (не хватает LBD или DBD) | А | KNR/ухо [ 50 ] (членистоногие) | NR0A1 | Кни | Zygotic Gap белок Jirps | КНЛ | |
| Беременный | DAX/SHP | NR0B1 | Dix1 | Чувствительное к дозировке реверс пола, критическая область гипоплазии надпочечников, на хромосоме X, ген 1 | NR0B1 | |||
| NR0B2 | Шп | Маленький Гетеродимер Партнер | NR0B2 | |||||
Из двух 0-фемилов 0A имеет семейный 1-подобный DBD, а 0B имеет уникальный LBD. Второй DBD Family 7, вероятно, связан с Family 1 DBD. Три, вероятно, семейство 1 NR из Biomphalaria glabrata обладают DBD вместе с LBD Family Family. [ 50 ] Размещение C. elegans NHR-1 ( Q21878 ) оспаривается: хотя большинство источников помещают его как NR1K1, [ 50 ] Ручная аннотация в Wormbase считает его членом NR2A. [ 63 ] Раньше была группа 2D, для которой единственным членом были Drosophila HR78/NR1D1 ( Q24142 ) и ортологи, но он был объединен в группу 2C позже из -за высокого сходства, образуя «группу 2C/D». [ 50 ] Нокаутные исследования мышей и фруктовых мух поддерживают такую объединенную группу. [ 64 ]
Эволюция
[ редактировать ]Тема дебатов была в отношении личности наследственного ядерного рецептора как лиганд-связывающего или сиротского рецептора . Эта дебаты начались более двадцати пяти лет назад, когда первые лиганды были идентифицированы как стероид млекопитающих и гормоны щитовидной железы. [ 65 ] Вскоре после этого идентификация рецептора экдизона у дрозофилы представила идею о том, что ядерные рецепторы были гормональными рецепторами, которые связывают лиганды с наномолярной аффинностью. В то время тремя известными лигандами ядерных рецепторов были стероиды, ретиноиды и гормон щитовидной железы, и из этих трех стероидов и ретиноидов были продукты метаболизма терпеноидов. Таким образом, было постулировано, что наследственный рецептор был бы связан молекулой терпеноидов. [ 66 ]
В 1992 году сравнение ДНК-связывающего домена всех известных ядерных рецепторов привело к построению филогенного дерева ядерного рецептора, которое указывало на то, что все ядерные рецепторы разделили общего предка. [ 67 ] В результате усилились усилия по обнаружению состояния первого ядерного рецептора, и к 1997 году была предложена альтернативная гипотеза: предков ядерный рецептор был рецептором сироты, и он приобрел лигандскую способность с течением времени. [ 7 ] Эта гипотеза была предложена на основе следующих аргументов:
- Последовательности ядерных рецепторов, которые были идентифицированы в самых ранних метазоанах (Cnidarians и Schistosoma ), были членами групп рецепторов Coup-TF, RXR и FTZ-F1. Как Coup-TF, так и FTZ-F1 являются рецепторами сирот, а RXR обнаруживается только лиганда только у позвоночных. [ 68 ]
- В то время как рецепторы сирот были известны гомологам членистоногих, не было идентифицировано никаких ортологов рецепторов с лигантами, которые не были идентифицированы за пределами позвоночных, что предполагает, что рецепторы-сирот старше, чем рецепторы. [ 69 ]
- Сиротаные рецепторы обнаруживаются среди всех шести подсемейств ядерных рецепторов, в то время как лиганд-зависимые рецепторы обнаруживаются среди трех. [ 7 ] Таким образом, поскольку лиганд-зависимые рецепторы считались преимущественно членом недавних подсемейств, казалось логичным, что они приобрели способность независимо связывать лиганды.
- Филогенетическое положение данного ядерного рецептора внутри дерева коррелирует с его ДНК-связывающим доменом и способностями димеризации, но не существует идентифицированной связи между лиганд-зависимым ядерным рецептором и химической природой его лиганда. В дополнение к этому, эволюционные отношения между лиганд-зависимыми рецепторами не имели особого смысла, так как тесно связанные рецепторы подсемейств, связанных лигандов, происходящих из совершенно разных путей биосинтеза (например, TRS и RARS). С другой стороны, подсемейства, которые не являются эволюционно связанными, связывают сходные лиганды (RAR и RXR связывают все транс-транс и 9-цис-ретиноевую кислоту соответственно). [ 69 ]
- В 1997 году было обнаружено, что ядерные рецепторы не существуют в статических условиях и при конформациях, но что лиганд может изменить равновесие между двумя состояниями. Кроме того, было обнаружено, что ядерные рецепторы могут регулировать лиганд независимым образом, либо посредством фосфорилирования, либо других посттрансляционных модификаций. Таким образом, это обеспечило механизм для того, как был регулируется наследственный рецептор сирот в зависимости от лиганда, и объяснил, почему консервативный домен связывания лиганда. [ 69 ]
В течение следующих 10 лет были проведены эксперименты, чтобы проверить эту гипотезу, и вскоре возникли контраргументы:
- Ядерные рецепторы были идентифицированы в недавно секвенированном геноме Demosponge Amphimedon Queenslandica , члена Porifera, самой древней филома метазоа. Геном A. Queenslandica содержит два ядерных рецептора, известные как AQNR1 и AQNR2, и оба были охарактеризованы для связывания и регулируемых лигандами. [ 70 ]
- Гомологи для лиганд-зависимых рецепторов позвоночных были обнаружены вне позвоночных у молсов и Platyhelminthes. Кроме того, было обнаружено, что ядерные рецепторы, обнаруженные у Cnidarians, имеют структурные лиганды у млекопитающих, которые могут отражать наследственную ситуацию.
- Два предполагаемых рецептора сирот , HNF4 и USP были обнаружены посредством анализа структурных и масс -спектрометрии для связывания жирных кислот и фосфолипидов соответственно. [ 56 ]
- Ядерные рецепторы и лиганды обнаруживаются намного менее специфичны, чем считались ранее. Ретиноиды могут связывать рецепторы млекопитающих, отличные от RAR и RXR, такие как, PPAR, RORB или Coup-TFII. Кроме того, RXR чувствителен к широкому диапазону молекул, включая ретиноиды, жирные кислоты и фосфолипиды. [ 71 ]
- Исследование эволюции стероидных рецепторов показало, что наследственный стероидный рецептор может связывать лиганд, эстрадиол. И наоборот, рецептор эстрогена, обнаруженный в моллюсках, является конститутивно активным и не связывал гормоны, связанные с эстрогенами. Таким образом, это дало пример того, как наследственный лиганд-зависимый рецептор может потерять свою способность связывать лиганды. [ 72 ]
Сочетание этого недавнего доказательства, а также углубленное исследование физической структуры связывающего домена ядерного рецептора привело к появлению новой гипотезы относительно наследственного состояния ядерного рецептора. Эта гипотеза предполагает, что наследственный рецептор может действовать как липидный датчик со способностью связывать, хотя и довольно слабо, несколько различных гидрофобных молекул, таких как ретиноиды, стероиды, гем и жирные кислоты. Благодаря его способности взаимодействовать с различными соединениями, этот рецептор посредством дупликаций либо потеряет свою способность к лиганд-зависимой активности, либо специализируется на очень специфическом рецепторе для конкретной молекулы. [ 71 ]
История
[ редактировать ]Ниже приведен краткий выбор ключевых событий в истории исследований ядерных рецепторов. [ 73 ]
- 1905 - Эрнест Старлинг придумал слово гормон
- 1926 - Эдвард Кальвин Кендалл и Тадес Рейхштейн изолировали и определили структуры кортизона и тироксина
- 1929 - Адольф Бутенандт и Эдвард Адельберт Дойси - независимо изолирован и определили структуру эстрогена
- 1958 - Элвуд Дженсен - изолировал рецептор эстрогена
- 1980 -е годы - Клонирование рецепторов эстрогена, глюкокортикоидов и гормонов щитовидной железы Пьером Шамбоном , Рональдом Эвансом и Бьорн Веннестрем соответственно
- 2004 - Пьер Шамбон, Рональд Эванс и Элвуд Дженсен были удостоены премии Альберта Ласкер за базовые медицинские исследования , награду, которая часто предшествует Нобелевской премии по медицине
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный PDB : 3E00 ; Чандра В., Хуан П., Хамуро Ю., Рагурам С., Ван Й., Беррис Т.П., Растинежад Ф. (ноябрь 2008 г.). «Структура интактного комплекса PPAR-гамма-RXR-ядерного рецептора на ДНК» . Природа . 456 (7220): 350–6. doi : 10.1038/nature07413 . PMC 2743566 . PMID 19043829 .
- ^ Эванс Р.М. (май 1988). «Сверхсемейство рецептора стероидов и гормонов щитовидной железы» . Наука . 240 (4854): 889–95. Bibcode : 1988sci ... 240..889e . doi : 10.1126/science.3283939 . PMC 6159881 . PMID 3283939 .
- ^ Олефски Дж. М. (октябрь 2001 г.). «Серия ядерного рецептора minireview» . Журнал биологической химии . 276 (40): 36863–4. doi : 10.1074/jbc.r100047200 . PMID 11459855 . S2CID 5497175 .
- ^ Jump up to: а беременный в Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schütz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, Evans RM (декабрь 1995 г.). «Суперсемейство ядерного рецептора: второе десятилетие» . Клетка . 83 (6): 835–9. doi : 10.1016/0092-8674 (95) 90199-X . PMC 6159888 . PMID 8521507 .
- ^ Jump up to: а беременный Novac N, Heinzel T (декабрь 2004 г.). «Ядерные рецепторы: обзор и классификация». Современные цели наркотиков. Воспаление и аллергия . 3 (4): 335–46. doi : 10.2174/1568010042634541 . PMID 15584884 .
- ^ Jump up to: а беременный в Комитет по номенклатуре ядерных рецепторов (апрель 1999 г.). «Объединенная система номенклатуры для суперсемейства ядерного рецептора» . Клетка . 97 (2): 161–3. doi : 10.1016/s0092-8674 (00) 80726-6 . PMID 10219237 . S2CID 36659104 .
- ^ Jump up to: а беременный в дюймовый Laudet V (декабрь 1997 г.). «Эволюция суперсемейства ядерных рецепторов: ранняя диверсификация от наследственного рецептора сирот». Журнал молекулярной эндокринологии . 19 (3): 207–26. doi : 10.1677/jme.0.0190207 . PMID 9460643 . S2CID 16419929 .
- ^ Escriva H, Langlois MC, Mendonça RL, Pierce R, Laudet V (май 1998). «Эволюция и диверсификация суперсемейства ядерного рецептора». Анналы нью -йоркской академии наук . 839 (1): 143–6. Bibcode : 1998nyasa.839..143e . doi : 10.1111/j.1749-6632.1998.tb10747.x . PMID 9629140 . S2CID 11164838 .
- ^ Рейцель А.М., Панг К., Райан Дж.Ф., Малликин Дж.С., Мартиндейл М.К., Баксеванис А.Д., Таррант А.М. (февраль 2011 г.). «Ядерные рецепторы из Ctenophore mnemiopsis leidyi отсутствуют ДНК-связывающего домена цинка: специфичная для линии потери или наследственное состояние при появлении суперсемейства ядерного рецептора?» Полем Эводево . 2 (1): 3. DOI : 10.1186/2041-9139-2-3 . PMC 3038971 . PMID 21291545 .
- ^ Bridgham JT, Eick GN, Larroux C, Deshpande K, Harms MJ, Gauthier ME, Ortlund EA, Degnan BM, Thornton JW (октябрь 2010 г.). «Эволюция белка с помощью молекулярного возиции: диверсификация суперсемейства ядерного рецептора от лиганд-зависимого предка» . PLOS Биология . 8 (10): E1000497. doi : 10.1371/journal.pbio.1000497 . PMC 2950128 . PMID 20957188 .
- ^ Sluder AE, Maina CV (апрель 2001 г.). «Ядерные рецепторы в нематодах: темы и вариации». Тенденции в генетике . 17 (4): 206–13. doi : 10.1016/s0168-9525 (01) 02242-9 . PMID 11275326 .
- ^ Читл Джарвела А.М., Пик Л (2017). «Функция и эволюция ядерных рецепторов в эмбриональном развитии насекомых». Современные темы в биологии развития . 125 : 39–70. doi : 10.1016/bs.ctdb.2017.01.003 . ISBN 9780128021729 Полем PMID 28527580 .
- ^ Schaaf MJ (2017). «Исследование ядерных рецепторов у рыбок данио» . Журнал молекулярной эндокринологии . 59 (1): R65 - R76. doi : 10.1530/jme-17-0031 . PMID 28438785 .
- ^ Jump up to: а беременный Zhang Z, Burch PE, Cooney AJ, Lanz RB, Pereira FA, Wu J, Gibbs RA, Weinstock G, Wheeler DA (апрель 2004 г.). «Геномный анализ семейства ядерных рецепторов: новое понимание структуры, регуляции и эволюции из генома крысы» . Исследование генома . 14 (4): 580–90. doi : 10.1101/gr.2160004 . PMC 383302 . PMID 15059999 .
- ^ Overington JP, Al-Lazikani B, Hopkins AL (декабрь 2006 г.). «Сколько существует целей наркотиков?». Природные обзоры. Открытие наркотиков . 5 (12): 993–6. doi : 10.1038/nrd2199 . PMID 17139284 . S2CID 11979420 .
- ^ Бенуа Г., Куни А., Гигур В., Ингрхам Х. , Лазар М., Маскат Г., Перлман Т., Рено Дж.П., Швабе Дж., Сладек Ф., Цай М.Дж. , Лаудет V (декабрь 2006 г.). «Международный союз фармакологии. LXVI. Ядерные рецепторы сироты». Фармакологические обзоры . 58 (4): 798–836. doi : 10.1124/pr.58.4.10 . PMID 17132856 . S2CID 2619263 .
- ^ Мохан Р., Хейман Р.А. (2003). «Модуляторы ядерных рецепторов сирот». Текущие темы в лекарственной химии . 3 (14): 1637–47. doi : 10.2174/1568026033451709 . PMID 14683519 .
- ^ Кумар Р., Томпсон Э.Б. (май 1999). «Структура рецепторов ядерного гормона». Стероиды . 64 (5): 310–9. doi : 10.1016/s0039-128x (99) 00014-8 . PMID 10406480 . S2CID 18333397 .
- ^ Klinge CM (май 2000). «Взаимодействие рецептора эстрогена с ко-активаторами и ко-репрессорами». Стероиды . 65 (5): 227–51. doi : 10.1016/s0039-128x (99) 00107-5 . PMID 10751636 . S2CID 41160722 .
- ^ Jump up to: а беременный Wärnmark A, Treuter E, Wright AP, Gustafsson JA (октябрь 2003 г.). «Функции активации 1 и 2 ядерных рецепторов: молекулярные стратегии для активации транскрипции» . Молекулярная эндокринология . 17 (10): 1901–9. doi : 10.1210/me.2002-0384 . PMID 12893880 . S2CID 31314461 .
- ^ Wu W, Loverde Pt (2021). «Идентификация и эволюция ядерных рецепторов у Platyhelmints» . Plos один . 16 (8): E0250750 (8): E0250750. Bibcode : 2021ploso..1650750W . doi : 10.1371/journal.pone.0250750 . PMC 8363021 . PMID 34388160 .
- ^ Weatherman RV, Fletterick RJ, Scanlan TS (1999). «Ядерные рецепторные лиганды и лиганд-связывающие домены». Ежегодный обзор биохимии . 68 : 559–81. doi : 10.1146/annurev.biochem.68.1.559 . PMID 10872460 .
- ^ Чандра В., Хуан П., Потлури Н., Ву Д., Ким Й., Растинежад Ф (март 2013 г.). «Многодоменная интеграция в структуре комплекса ядерного рецептора HNF-4α» . Природа . 495 (7441): 394–8. Bibcode : 2013natur.495..394c . doi : 10.1038/nature11966 . PMC 3606643 . PMID 23485969 .
- ^ Лу Х, Торессон Г., Бенод С., Су Дж.Х., Филипс К.Дж., Уэбб П., Густафссон Дж.А. (март 2014 г.). «Структура рецептора рецептора ретиноида x α-ливер X β (RXRα-LXRβ) на ДНК». Природа структурная и молекулярная биология . 21 (3): 277–81. doi : 10.1038/nsmb.2778 . PMID 24561505 . S2CID 23226682 .
- ^ ВВП : 2C7A ; Roemer SC, Donham DC, Sherman L, Pon VH, Edwards DP, Churchill ME (декабрь 2006 г.). «Структура комплекса прогестерона рецептор-дезоксирибонуклеиновой кислоты: новые взаимодействия, необходимые для связывания с полусменными элементами ответа» . Молекулярная эндокринология . 20 (12): 3042–52. doi : 10.1210/me.2005-0511 . PMC 2532839 . PMID 16931575 .
- ^ PDB : 3L0L ; Джин Л., Мартиновский Д., Чжэн С., Вада Т, Се В, Ли Ю (май 2010 г.). «Структурные основы для гидроксихолестерина как природных лигандов ядерного рецептора сироты Роргамма» . Молекулярная эндокринология . 24 (5): 923–9. doi : 10.1210/me.2009-0507 . PMC 2870936 . PMID 20203100 .
- ^ Jump up to: а беременный в дюймовый Amoutzias GD, Pichler EE, Mian N, De Graaf D, Imsiridou A, Robinson-Rechavi M, Bornberg-Bauer E, Robertson DL, Oliver SG (июль 2007 г.). «Атлас взаимодействия белка для ядерных рецепторов: свойства и качество димеризационной сети на основе концентраторов» . BMC Системная биология . 1 : 34. DOI : 10.1186/1752-0509-1-34 . PMC 1971058 . PMID 17672894 .
- ^ Line MJ, Porkka KP, Kang Z, Savinainen KJ, Jänne Oa, Tammela TL, Vessella RL, Palvimo JJ, Visakorpi T (февраль 2004 г.). «Экспрессия коррегуляторов рецептора андрогена при раке простаты». Клиническое исследование рака . 10 (3): 1032-40. Doi : 10.1158/1078-0432.ccr-0990-3 . PMID 14871982 . S2CID 8038717 .
- ^ Klinge CM, Bodenner DL, Desai D, Niles RM, Traish AM (май 1997). «Связывание ядерных рецепторов типа II и рецептора эстрогена с полным и полусединным элементом ответа эстрогена in vitro» . Исследование нуклеиновых кислот . 25 (10): 1903–12. doi : 10.1093/nar/25.10.1903 . PMC 146682 . PMID 9115356 .
- ^ Rual, Жан-Франсуа; Венкатесан, Кавита; Хао, Тонг; Хирозане-Кишикава, Томоко; Дрикот, Амели; Оболочка; Берриз, Габриэль Ф.; Гиббонс, Фрэнсис Д.; Дрез, Матиджа; Ayivi-guedehoussou, nono; Klitgord, Niels (2005-10-20). «На пути к масштабе протеома карты взаимодействия белкового белка человека» . Природа . 437 (7062): 1173–1178. Bibcode : 2005natur.437.1173r . doi : 10.1038/nature04209 . ISSN 1476-4687 . PMID 16189514 . S2CID 4427026 .
- ^ Альберс, Майкл; Кранц, Харальд; Кобер, Инго; Кайзер, Кармен; Клинк, Мартин; Suckow, Jörg; Керн, Рейнер; Koegl, Manfred (февраль 2005 г.). «Автоматизированный двухгибридный скрининг для белков-интерезации ядерного рецептора» . Молекулярная и клеточная протеомика . 4 (2): 205–213. doi : 10.1074/mcp.m400169-mcp200 . ISSN 1535-9476 . PMID 15604093 . S2CID 14876486 .
- ^ McKenna NJ, Lanz RB, O'Malley BW (июнь 1999 г.). «Ядерные рецепторы Coregulators: клеточная и молекулярная биология» . Эндокринные обзоры . 20 (3): 321–344. doi : 10.1210/edrv.20.3.0366 . PMID 10368774 . S2CID 10182146 .
- ^ Glass CK, Rosenfeld MG (январь 2000 г.). «Обмен ядерных рецепторов в области транскрипции ядерных рецепторов» . Гены и развитие . 14 (2): 121–41. doi : 10.1101/gad.14.2.121 . PMID 10652267 . S2CID 12793980 .
- ^ Аранда А, Паскуал А (июль 2001 г.). «Рецепторы ядерного гормона и экспрессия генов». Физиологические обзоры . 81 (3): 1269–304. doi : 10.1152/physrev.2001.81.3.1269 . HDL : 10261/79944 . PMID 11427696 . S2CID 5972234 .
- ^ Копленд JA, Шеффилд-Мур М., Колдич-Зиванович Н., Джентри С., Лампро Г., Цорцату-Статопулу Ф., Зумпурлис В., Урбан Р.Дж., Влахопулос С.А. (июнь 2009 г.). «Половые стероидные рецепторы при дифференцировке скелета и эпителиальной неоплазии: возможно ли тканевое вмешательство?». Биологии . 31 (6): 629–41. doi : 10.1002/bies.200800138 . PMID 19382224 . S2CID 205469320 .
- ^ Brzozowski AM, Pike AC, Dauter Z, Hubbard RE, Bonn T, Engström O, Ohman L, Greene GL, Gustafsson JA, Carlquist M (октябрь 1997 г.). «Молекулярная основа агонизма и антагонизма в рецепторе эстрогена». Природа . 389 (6652): 753–8. Bibcode : 1997natur.389..753b . doi : 10.1038/39645 . PMID 9338790 . S2CID 4430999 .
- ^ Шиау А.К., Барстад Д., Лория П.М., Ченг Л., Кушнер П.Дж., Агард Д.А., Грин Г.Л. (декабрь 1998 г.). «Структурная основа распознавания рецептора эстрогена/коактиватора и антагонизма этого взаимодействия тамоксифеном» . Клетка . 95 (7): 927–37. doi : 10.1016/s0092-8674 (00) 81717-1 . PMID 9875847 . S2CID 10265320 .
- ^ Gronemeyer H, Gustafsson JA, Laudet V (ноябрь 2004 г.). «Принципы модуляции суперсемейства ядерного рецептора». Природные обзоры. Открытие наркотиков . 3 (11): 950–64. doi : 10.1038/nrd1551 . PMID 15520817 . S2CID 205475111 .
- ^ Busch BB, Stevens WC, Martin R, Ordentlich P, Zhou S, Sapp DW, Horlick RA, Mohan R (ноябрь 2004 г.). «Идентификация селективного обратного агониста для ядерного рецептора-рецептора ядерного рецептора альфа-рецептора, связанного с рецептором». Журнал лекарственной химии . 47 (23): 5593–6. doi : 10.1021/jm049334f . PMID 15509154 .
- ^ Смит К.Л., О'Мэлли Б.В. (февраль 2004 г.). «Функция Coregulator: ключ к пониманию тканевой специфичности селективных модуляторов рецепторов» . Эндокринные обзоры . 25 (1): 45–71. doi : 10.1210/er.2003-0023 . PMID 14769827 .
- ^ Pascual G, Glass CK (октябрь 2006 г.). «Ядерные рецепторы по сравнению с воспалением: механизмы трансрепрессии». Тенденции в эндокринологии и метаболизме . 17 (8): 321–7. doi : 10.1016/j.tem.2006.08.005 . PMID 16942889 . S2CID 19612552 .
- ^ Björnström L, Sjöberg M (июнь 2004 г.). «Эстроген-рецептор-зависимая активация AP-1 посредством негеномной передачи сигналов» . Ядерный рецептор . 2 (1): 3. DOI : 10.1186/1478-1336-2-3 . PMC 434532 . PMID 15196329 .
- ^ Zivadinovic D, Gametchu B, Watson CS (2005). «Уровни мембранного рецептора эстрогена-альфа в клетках рака молочной железы MCF-7 предсказывают реакции цАМФ и пролиферации» . Исследование рака молочной железы . 7 (1): R101–12. doi : 10.1186/bcr958 . PMC 1064104 . PMID 15642158 .
- ^ Kousteni S, Bellido T, Plotkin Li, O'Brien CA, Bodenner DL, Han L, Han K, Digregorio GB, Katzenellenbogen JA, Katzenellenbogen BS, Roberson PK , Weinstein RS, Jilka RL, Manolagas SC (март 2001 г.). «Негенотропная, неспецифическая для полов передачи сигналов через рецепторы эстрогена или андрогена: диссоциация от транскрипционной активности» . Клетка . 104 (5): 719–30. doi : 10.1016/s0092-8674 (01) 00268-9 . PMID 11257226 . S2CID 10642274 .
- ^ Storey NM, Gentile S, Ullah H, Russo A, Muessel M, Erxleben C, Armstrong DL (март 2006 г.). «Быстрая передача сигналов на плазматической мембране ядерным рецептором для гормона щитовидной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (13): 5197–201. Bibcode : 2006pnas..103.5197s . doi : 10.1073/pnas.0600089103 . PMC 1458817 . PMID 16549781 .
- ^ Стори Н.М., О'Брайан Дж.П., Армстронг Д.Л. (январь 2002 г.). «RAC и RHO опосредуют противодействие гормональной регуляции калиевого канала, связанного с эфиром, калиевого калия» . Текущая биология . 12 (1): 27–33. Bibcode : 2002cbio ... 12 ... 27 с . doi : 10.1016/s0960-9822 (01) 00625-x . PMID 11790300 . S2CID 8608805 .
- ^ Jump up to: а беременный Мартин Н.П., Маррон Фернандес де Веласко Е., Мизуно Ф., Скаппини Э.Л., Глосс Б., Эркслбен С., Уильямс Дж.Г., Стэплтон Х.М., Джентиле С., Армстронг Д.Л. (сентябрь 2014 г.). «Быстрый цитоплазматический механизм регуляции PI3 -киназы с помощью ядерного рецептора гормона щитовидной железы, TRβ и генетических доказательств его роли в созревании синапсов гиппокампа мыши in vivo» . Эндокринология . 155 (9): 3713–24. doi : 10.1210/en.2013-2058 . PMC 4138568 . PMID 24932806 .
- ^ Гилберт меня (январь 2004 г.). «Изменения в синаптической передаче и пластичности в области CA1 взрослого гиппокампа после гипотиреоза развития» . Исследование мозга. Исследование мозга развития . 148 (1): 11–8. doi : 10.1016/j.devbrainres.2003.09.018 . PMID 14757514 .
- ^ Moeller LC, Broecker-Preuss M (август 2011 г.). «Транскрипционная регуляция путем неклассического действия гормона щитовидной железы» . Исследование щитовидной железы . 4 (Suppl 1): S6. doi : 10.1186/1756-6614-4-S1-S6 . PMC 3155112 . PMID 21835053 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Kaur S, Jobling S, Jones CS, Noble LR, Routledge EJ, Lockyer AE (7 апреля 2015 г.). «Ядерные рецепторы Biomphalaria glabrata и Lottia gigantea: последствия для разработки новых модельных организмов» . Plos один . 10 (4): E0121259. BIBCODE : 2015PLOSO..1021259K . doi : 10.1371/journal.pone.0121259 . PMC 4388693 . PMID 25849443 .
- ^ Burris TP, De Vera IM, Cote I, Flaveny CA, Wanninayake US, Chatterjee A, Walker JK, Steinauer N, Zhang J, Coons LA, Korach KS, Cain DW, Hollenberg AN, Webb P, Forrest D, Jetten Am, Edwards Д.П., Гримм С.Л., Хартиг С., Ланге К.А., Ришер Дж.К., Сарториус К.А., Тетель М., Биллон С., Элгенди Б., Хегази Л., Гриффетт К., Пейнетти Н., Бернштейн К.Л., Хьюз Т.С., Ситаула С., Стейльрак К.Р., Калвер А, Murray MH, Finck BN, Cidlowski JA (ноябрь 2023 г.). Ohlstein E (ред.). «Международный союз базовой и клинической фармакологии CXIII: суперсемейство ядерного рецептора - AUPDATE 2023» . Фармакологические обзоры . 75 (6): 1233–1318. doi : 10.1124/pharmrev.121.000436 . ISSN 0031-6997 . PMC 10595025 . PMID 37586884 .
- ^ Crossgrove K, Laudet V, Maina CV (февраль 2002 г.). «Dirofilaria Immitis кодирует DI-NHR-7, толчок гена E78, регулируемого Drorophila Ecdysone» Молекулярная и биохимическая паразитология 119 (2): 169–7 Doi : 10.1016/s0166-6851 (01) 00412-1 11814569PMID
- ^ "Sex-1 (ген)" . Wormbase: нематодный информационный ресурс .
- ^ Otte K, Kranz H, Kober I, Thompson P, Hoefer M, Haubold B, Remmel B, Voss H, Kaiser C, Albers M, Cheruvallath Z, Jackson D, Casari G, Koegl M, Pääbo S, Mous J, Kremoser C , Deuschle U (февраль 2003 г.). «Идентификация фарнезоидного X -рецептора бета в качестве нового ядерного рецептора млекопитающего, воспринимающего ланостерол» . Молекулярная и клеточная биология . 23 (3): 864–72. doi : 10.1128/mcb.23.3.3.864-872.2003 . PMC 140718 . PMID 12529392 .
- ^ «Отчет гена Flybase: DMEL \ HR96» . Полетная база . Получено 14 августа 2019 года .
- ^ Jump up to: а беременный Schwabe JW, Teichmann SA (январь 2004 г.). «Ядерные рецепторы: эволюция разнообразия». Science's Stke . 2004 (217): PE4. doi : 10.1126/stke.2172004pe4 . PMID 14747695 . S2CID 20835274 .
- ^ Kruse SW, Suino-Powell K, Zhou Xe, Kretchman JE, Reynolds R, Vonrhein C, et al. (Сентябрь 2008 г.). «Идентификация ядерного рецептора Orphan Coup-TFII в качестве рецептора, активируемого ретиноевой кислотой» . PLOS Биология . 6 (9): E227. doi : 10.1371/journal.pbio.0060227 . PMC 2535662 . PMID 18798693 .
- ^ Im, Bonneton F, Moras D, et. (Март 2017). Антеровой силой является Ориентар . Наука достижения 3 (3): E1 Bibcode : 2017scia .... 3E178M doi : 10.1126/sciadv . PMC 5375646 . PMID 28435861 .
- ^ Jump up to: а беременный Khalturin K, Billas I, Chebaro Y, Reitzel AM, Tarrant AM, Laudet V, Markov GV (ноябрь 2018). «Рецепторы NR3E у Cnidarians: новое семейство родственников стероидных рецепторов расширяет возможные механизмы для связывания лиганда» . J Стероидный биохим Mol Biol . 184 : 11–19. doi : 10.1016/j.jsbmb.2018.06.014 . PMC 6240368 . PMID 29940311 .
- ^ Wu, W, Loverde Pt (сентябрь 2023 г.). «Обновленные знания и предлагаемая номенклатура для ядерных рецепторов с двумя доменами связывания ДНК (2DBD-NRS)» . Plos один . 18 (9): E0286107. Bibcode : 2023ploso..1886107W . doi : 10.1371/journal.pone.0286107 . PMC 10497141 . PMID 37699039 .
- ^ Wu W, Niles EG, Hirai H, Loverde Pt (февраль 2007 г.). «Эволюция новой подсемейства ядерных рецепторов с членами, каждый из которых содержат два домена связывания ДНК» . BMC Evol Biol . 7 (27): 27. Bibcode : 2007bmcee ... 7 ... 27w . doi : 10.1186/1471-2148-7-27 . PMC 1810520 . PMID 17319953 .
- ^ Huang W, Xu F, Li J, Li L, Que H, Zhang G (август 2015 г.). «Эволюция новой подсемейства ядерных рецепторов с акцентом на члена из тихоокеанского устричного гигас». Ген . 567 (2): 164–72. doi : 10.1016/j.gene.2015.04.082 . PMID 25956376 .
- ^ «NHR-1 (ген)» . Wormbase: нематодный информационный ресурс .
- ^ Marxreiter S, Thummel CS (февраль 2018 г.). «Взрослые функции для ядерного рецептора Drosophila DHR78» . Динамика развития . 247 (2): 315–322. doi : 10.1002/dvdy.24608 . PMC 5771960 . PMID 29171103 .
- ^ Эванс Р.М. (май 1988). «Сверхсемейство рецептора стероидов и гормонов щитовидной железы» . Наука . 240 (4854): 889–95. Bibcode : 1988sci ... 240..889e . doi : 10.1126/science.3283939 . PMC 6159881 . PMID 3283939 .
- ^ Мур Д.Д. (январь 1990). «Разнообразие и единство в рецепторах ядерного гормона: суперсемейство терпеноидного рецептора». Новый биолог . 2 (1): 100–5. PMID 1964083 .
- ^ Laudet V, Hänni C, Coll J, Catzeflis F, Stéhelin D (март 1992 г.). «Эволюция суперсемейства гена ядерного рецептора» . Embo Journal . 11 (3): 1003–13. doi : 10.1002/j.1460-2075.1992.tb05139.x . PMC 556541 . PMID 1312460 .
- ^ Escriva H, Safi R, Hänni C, Langlois MC, Saumitou-Laprade P, Stehelin D, Capron A, Pierce R, Laudet V (июнь 1997 г.). «Связывание лигандов было приобретено во время эволюции ядерных рецепторов» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (13): 6803–8. Bibcode : 1997pnas ... 94.6803e . doi : 10.1073/pnas.94.13.6803 . PMC 21239 . PMID 9192646 .
- ^ Jump up to: а беременный в Escriva H, Delaunay F, Laudet V (август 2000 г.). «Связывание лигандов и эволюция ядерного рецептора». Биологии . 22 (8): 717–27. doi : 10.1002/1521-1878 (200008) 22: 8 <717 :: AID-bies5> 3.0.co; 2-I . PMID 10918302 . S2CID 45891497 .
- ^ Bridgham JT, Eick GN, Larroux C, Deshpande K, Harms MJ, Gauthier ME, Ortlund EA, Degnan BM, Thornton JW (октябрь 2010 г.). «Эволюция белка с помощью молекулярного возиции: диверсификация суперсемейства ядерного рецептора от лиганд-зависимого предка» . PLOS Биология . 8 (10): E1000497. doi : 10.1371/journal.pbio.1000497 . PMC 2950128 . PMID 20957188 .
- ^ Jump up to: а беременный Марков Гв, Laudet V (март 2011 г.). «Происхождение и эволюция лиганд-связывающей способности ядерных рецепторов». Молекулярная и клеточная эндокринология . Эволюция рецепторов ядерного гормона. 334 (1–2): 21–30. doi : 10.1016/j.mce.2010.10.017 . PMID 21055443 . S2CID 33537979 .
- ^ Thornton JW, нуждается в E, экипажах D (сентябрь 2003 г.). «Воскресение наследственного стероидного рецептора: древнее происхождение передачи сигналов эстрогена». Наука . 301 (5640): 1714–7. Bibcode : 2003sci ... 301.1714t . doi : 10.1126/science.1086185 . PMID 14500980 . S2CID 37628350 .
- ^ Тата -младший (июнь 2005 г.). «Сто лет гормонов» . Embo сообщает . 6 (6): 490–6. doi : 10.1038/sj.embor.7400444 . PMC 1369102 . PMID 15940278 .
Внешние ссылки
[ редактировать ]- Ядерные+рецепторы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Винсент Лаудет (2006). «Сборник Iuphar фармакологии и классификации суперсемейства ядерного рецептора 2006E» . Компендиум ядерного рецептора . Международный союз базовой и клинической фармакологии. Архивировано с оригинала 2015-04-02 . Получено 2008-02-21 .
- « ядерного рецептора Онлайн -журнал » . Домашняя страница . Опубликовано BioMed Central (больше не принимая заявки с мая 2007 года) . Получено 2008-02-21 .
- «Ресурс ядерного рецептора» . Джорджтаунский университет. Архивировано из оригинала 2008-05-11 . Получено 2008-02-21 .
- «Атлас передачи сигналов ядерного рецептора (рецепторы, коактиваторы, корепрессоры и лиганды)» . Консорциум NURSA . Получено 2008-02-21 .
исследовательский консорциум и база данных, финансируемый NIH; Включает журнал с открытым доступом PubMed, индексированную журнал, передача сигналов ядерного рецептора
- «Ресурс ядерного рецептора» . Джек Ванден Хевел . Получено 2009-09-21 .