Ариловой углеводородной рецептор
| Ах | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | Ahr , Bhlhe76, арилоглеводородный рецептор, рецептор ариловой углеводороды, RP85 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 600253 ; MGI : 105043 ; Гомологен : 1224 ; GeneCards : Ahr ; OMA : AHR - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Рецептор ариловой углеводороды (также известный как AHR , AHR , AHR , AHR , AH -рецептор или в качестве рецептора диоксина ) является белком , который у людей кодируется геном AHR . Рецептор ариловой углеводороды является транскрипционным фактором , который регулирует экспрессию генов. Первоначально считалось, что он функционирует главным образом как датчик ксенобиотических химических веществ, а также как регулятор ферментов, таких как цитохром P450 , которые метаболизируют эти химические вещества. Наиболее заметными из этих ксенобиотических химических веществ являются ароматические (арильные) углеводороды, из которых рецептор получает свое название.
Совсем недавно было обнаружено, что AHR активируется (или деактивируется) рядом эндогенных производных индола , таких как кинуренин . В дополнение к регуляции ферментов метаболизма, AHR играет роль в регуляции иммунных клеток, поддержании стволовых клеток и клеточной дифференцировке . [ 5 ] [ 6 ] [ 7 ]
Ариловой углеводородной рецептор является членом семейства спиральной спирали основных факторов транскрипции . AHR связывает несколько экзогенных лигандов, таких как природные флавоноиды растений , полифенолы и индолы , а также синтетические полициклические ароматические углеводороды и диоксин-подобные соединения . AHR является цитозольным транскрипционным фактором, который обычно неактивен, связанный с несколькими ко-чаперонами . При связывании лиганда такими как 2,3,7,8-тетрахлордибензо- p -диоксин (TCDD), патроны диссоциируют , что приводит к транслокации AHR в ядро и димеризацию с помощью ARNT ( AHR-ядерный транслокатор ) , что приводит к изменениям гена с химическими веществами , Полем
Протеиновые функциональные домены
[ редактировать ]
AHR Белок содержит несколько доменов, критических для функции, и классифицируется как член основного семейства спирали-петли-спираль / per-arnt-sim (BHLH / PAS) факторов транскрипции . [ 8 ] [ 9 ] Мотив BHLH расположен в N-конце белка и является обычным сущностью в различных факторах транскрипции . [ 10 ] Члены суперсемейства BHLH имеют две функционально отличительные и высоко консервативные области. Первый-это основная региона (B), которая участвует в связывании фактора транскрипции с ДНК . Вторым является область спиральной петли-спирали (HLH), которая облегчает взаимодействие белка белка. Также с AHR содержатся два домена PAS, PAS-A и PAS-B, которые представляют собой участки 200–350 аминокислот , которые демонстрируют гомологию высокой последовательности для белковых доменов, которые первоначально были обнаружены в периоде генов Drosophila (PER) и и и и целеустремленные (SIM) и в партнере AHR по димеризации ариловой углеводородной рецептор ядерного транслокатора (ARNT). [ 11 ] Домены PAS поддерживают специфические вторичные взаимодействия с другими доменом PAS, содержащими белки, как в случае AHR и ARNT, так что могут образовываться димерные и гетеромерные белковые комплексы. Сайт связывания лиганда AHR содержится в домене PAS-B [ 12 ] и содержит несколько консервативных остатков, критических для связывания лиганда. [ 13 ] Наконец, домен, богатый глутамином (Q-Rich), расположен в С-концевой области белка и участвует в рекрутировании и трансактивации ко-активатора. [ 14 ]
Лиганды
[ редактировать ]AHR -лиганды, как правило, были классифицированы на две категории, синтетические или природные. Первыми лигандами, которые были обнаружены, были синтетические ароматические углеводороды, такие как дибензодиоксины , дибензофураны , бифенилы и нехлорированные нафтофлавоны наряду с природными полициклическими арокартонами ( 3-метилхолантрон , бензой [ а ] пиранция полихлорированные . [ 15 ] [ 16 ] Ряд синтетических лигандов был разработан в качестве потенциального лечения рака молочной железы. [ 17 ]
Исследования были сосредоточены на других природных соединениях с надеждой идентификации эндогенного лиганда. Природные соединения, которые были идентифицированы как лиганды AHR, включают производные триптофана , такие как краситель индиго и индирубин , [ 18 ] тетрапирролы, такие как билирубин , [ 19 ] арахидоновой кислоты метаболиты липоксин A4 и простагландин G , [ 20 ] Модифицированный липопротеин с низкой плотностью [ 21 ] и несколько диетических каротиноидов . [ 16 ] Одним из предположений, сделанных в поиске эндогенного лиганда, является то, что лиганд будет рецепторным агонистом . Однако работа Savouret et al. показал, что это может быть не так, поскольку их результаты демонстрируют, что 7-кетохолестерин конкурентно ингибирует трансдукцию сигнала AHR. [ 22 ]
Carbidopa - это селективный модулятор рецептора ариловой углеводороды (SAHRM). [ 23 ] Другие SAHRM включают микробные 1,4-дигидрокси-2-нафтоевая кислота [ 24 ] и растение 3,3'-диинтолилметан. [ 25 ]
Индолокарбазол [ нужно разъяснения ] (ICZ) является одним из самых сильных негалогенированных агонистов для AHR in vitro. [ 26 ]
Лиганд-независимая активность AHR можно увидеть в AHR млекопитающих. AHR млекопитающего не нуждается в экзогенной лиганд-зависимой активации, чтобы быть функциональной, и это, по-видимому, имеет место для его роли в регуляции экспрессии некоторых изоформ трансформирующего фактора роста (TGF-B) . Это не означает, что лиганд-зависимая активация AHR не требуется для функционирования AHR в этих случаях, но что, если лиганд необходим, он предоставляется эндогенно по рассматриваемым клеткам или тканям, и его идентичность неизвестна. [ 27 ]
Сигнальный путь
[ редактировать ]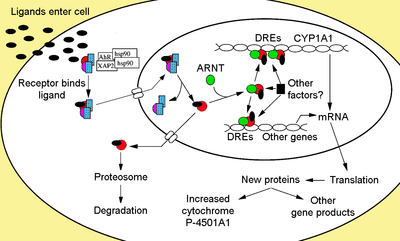

Цитозольный комплекс
[ редактировать ]Не связанный с лиганд AHR сохраняется в цитоплазме как неактивный белковый комплекс, состоящий из димера HSP90 , [ 28 ] [ 29 ] Простагландин Есинтаза 3 (Ptges3, P23) [ 30 ] [ 31 ] [ 32 ] [ 33 ] и единая молекула белка иммунофилиноподобному , подобного белкам , также известному как вирус Hpatitis B-вирус, x-ассоциированный белок 2 (XAP2), [ 34 ] AHR взаимодействующий белок ( AIP ), [ 35 ] [ 36 ] и AHR-активированный 9 (ARA9). [ 37 ] Дисмер HSP90, наряду с PTGES3 (P23), играет многофункциональную роль в защите рецептора от протеолиза, ограничивая рецептор в конформационном рецептивном рецептивном рецепте с связыванием лиганда и предотвращению преждевременного связывания ARNT . [ 12 ] [ 31 ] [ 33 ] [ 38 ] [ 39 ] [ 40 ] AIP взаимодействует с карбоксил-терминалом HSP90 и связывается с последовательности ядерной локализации AHR (NLS), предотвращая неуместный перевозку рецептора в ядро. [ 41 ] [ 42 ] [ 43 ]
Сигнальный путь трансформирующего фактора роста (TGF-β)
[ редактировать ]Цитокины TGF-β являются членами семейства сигнальных белков, который включает в себя активин, узловое подсемейство, морфогенетические белки кости, коэффициенты роста и дифференцировки и подсемейство ингибитора Мюллера. Передача сигналов TGF-β играет важную роль в физиологии и развитии клеток, ингибируя пролиферацию клеток, способствуя апоптозу, вызывая дифференцировку и определяя судьбу развития у позвоночных и беспозвоночных. [ 44 ] Активаторы TGF-β включают протеазы, такие как плазмин, катепсины и кальпаины. Тромбоспондин 1, гликопротеин, который ингибирует ангиогенез, и матриксная металлопротеиназа 2 (MMP-2). Сам внеклеточный матрикс, по-видимому, играет важную регуляторную роль в передаче сигналов TGF-β. [ 45 ] [ 46 ]
Активация рецептора
[ редактировать ]При связывании лиганда с AHR выпускается AIP, что приводит к воздействию NLS, который находится в области BHLH, [ 47 ] приводя к импорту в ядро. [ 48 ] Предполагается, что, как только в ядре, HSP90 диссоциирует, вызывая два домена PAS, позволяющие связывать ARNT. [ 40 ] [ 49 ] [ 50 ] [ 51 ] Активированный комплекс гетеродимера AHR/ARNT затем способен прямо или косвенно взаимодействовать с ДНК путем связывания с последовательностями распознавания, расположенных в 5'-регуляторной области генов, чувствительных к диоксинам. [ 40 ] [ 50 ] [ 52 ]
Связывание ДНК (элемент ксенобиотического ответа - XRE)
[ редактировать ]Классический мотив распознавания комплекса AHR/ARNT, называемый элементом AHR-, диоксин- или ксенобиотиков (AHRE, DRE или XRE), содержит основную последовательность 5'-GCGTG-3 ' [ 53 ] В рамках консенсусной последовательности 5'-t/gngcgtga/cg/ca-3 ' [ 54 ] [ 55 ] В промоторной области генов, реагирующих на AHR. Гетеродимер AHR/ARNT напрямую связывает последовательность ядра AHRE/DRE/XRE асимметричным образом, так что ARNT связывает 5'-GTG-3 'и связывание AHR 5'-TC/TGC-3'. [ 56 ] [ 57 ] [ 58 ] Недавние исследования показывают, что второй тип элемента, называемый AHRE-II, 5'-CATG (N6) C [T/A] TG-3 ', способен косвенно действовать с комплексом AHR/ARNT. [ 59 ] [ 60 ] Независимо от элемента ответа, результатом является различные дифференциальные изменения в экспрессии генов.
Функциональная роль в физиологии и токсикологии
[ редактировать ]Роль в разработке
[ редактировать ]С точки зрения эволюции, самая старая физиологическая роль AHR находится в развитии. Предполагается, что AHR развивался от беспозвоночных , где он служил независимой от лиганда роли в нормальных процессах развития. [ 61 ] Гомолог AHR у Drosophila , бесцветный (SS) необходим для развития дистальных сегментов антенны и ноги. [ 62 ] [ 63 ] SS димеризуется с танго (TGO), который является гомологом ARNT млекопитающего, чтобы инициировать транскрипцию генов. Эволюция рецептора у позвоночных приводила к способности связывать лиганды и, возможно, помогла людям развиваться до переноса дымных пожаров. В развивающихся позвоночных AHR, по -видимому, играет роль в клеточной пролиферации и дифференцировке. [ 64 ] Несмотря на отсутствие четкого эндогенного лиганда, AHR, по -видимому, играет роль в дифференциации многих путей развития, включая гематопоэс, [ 65 ] лимфоидные системы, [ 66 ] [ 67 ] Т-клетки, [ 68 ] нейроны, [ 69 ] и гепатоциты. [ 70 ] Также было обнаружено, что AHR имеет важную функцию в гемопоэтических стволовых клетках: антагонизм AHR способствует их самообновлению и экспансии Ex-Vivo [ 71 ] и участвует в дифференциации мегакариоцитов. [ 72 ] Во взрослом возрасте сигнализация связана с реакцией на стресс, а мутации в AHR связаны с серьезным депрессивным расстройством. [ 73 ]
Адаптивный и врожденный ответ
[ редактировать ]Адаптивный ответ проявляется как индукция ксенобиотических метаболизирующих ферментов. Свидетельство этого ответа впервые наблюдалось из индукции цитохрома P450, семейства 1, подсемейства A, полипептида 1 (CYP1A1), полученного в результате воздействия TCDD, которое было определено как непосредственно связано с активацией пути сигнализации AHR. [ 74 ] [ 75 ] [ 76 ] Поиск других метаболизирующих генов, вызванных лигандами AHR, из -за присутствия DRES привел к идентификации «батареи гена AHR», метаболизирующих ферменты I и фазы II, состоящие из CYP1A1 , CYP1A2 , CYP1B1 , NQO1, ALDH3A1,. UGT1A2 и GSTA1. [ 77 ] Предположительно, позвоночные имеют эту функцию, чтобы иметь возможность обнаружить широкий спектр химических веществ, что указывает широкий спектр субстратов AHR, способных связывать и облегчить их биотрансформацию и устранение. AHR может также сигнализировать о наличии токсичных химических веществ в пище и вызвать отвращение к таким продуктам. [ 78 ]
Активация AHR, по -видимому, также важна для иммунологических реакций и ингибирования воспаления [ 67 ] Благодаря активации интерлейкина 22 [ 79 ] и подавление ответа Th17 . [ 80 ] Нокдаун AHR в основном подавляет экспрессию генов врожденного иммунитета в клетках THP-1 . [ 81 ]
Токсичный ответ
[ редактировать ]Расширение адаптивного ответа являются токсическими реакциями, вызванными активацией AHR. Токсичность является результатом двух разных способов передачи сигналов AHR. Первый - это побочный эффект адаптивного ответа, в котором индукция метаболизирующих ферментов приводит к выработке токсичных метаболитов. Например, полициклический ароматический углеводородный бензо [ A ] пирен (BAP), лиганд для AHR, вызывает собственный метаболизм и биоактивацию для токсического метаболита посредством индукции CYP1A1 и CYP1B1 в нескольких тканях. [ 82 ] Второй подход к токсичности является результатом аберрантных изменений в глобальной транскрипции генов, помимо тех, которые наблюдаются в «батареи гена AHR». Эти глобальные изменения в экспрессии генов приводят к неблагоприятным изменениям в клеточных процессах и функции. [ 83 ] Анализ микрочипов оказался наиболее полезным в понимании и характеристике этого ответа. [ 64 ] [ 84 ] [ 85 ] [ 86 ]
Ксенобиотические метаболизирующие ферменты помогают с метаболическим процессом путем трансформации и экскреции химических веществ. Наиболее мощным индуктором CYP1A1 является 2,3,7,8-тетрахлордибензо-p-диоксин (TCDD). Кроме того, TCDD индуцирует широкий спектр биохимических и токсических эффектов, таких как тератогенез, иммуносупрессия и продвижение опухоли. Известно, что большинство, если не все, эффекты, вызванные TCDD и другими ПАУ, опосредованы AHR, который имеет высокую аффинность связывания с TCDD. [ 44 ]
Белковое взаимодействие
[ редактировать ]В дополнение к белковым взаимодействиям, упомянутым выше, было также показано, что AHR взаимодействует со следующим:
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000106546 - Ensembl , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000019256 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Эссер C (2016). «Ариловый углеводородный рецептор в иммунитете: инструменты и потенциал». Подавление и регуляция иммунных ответов . Методы в молекулярной биологии. Тол. 1371. С. 239–57. doi : 10.1007/978-1-4939-3139-2_16 . ISBN 978-1-4939-3138-5 Полем PMID 26530806 .
- ^ Kawajiri K, Fujii-Kuriyama Y (май 2017). «Рецептор арилглетодорода: многофункциональный химический датчик для защиты хозяина и гомеостатического поддержания» . Экспериментальные животные . 66 (2): 75–89. doi : 10.1538/expanim.16-0092 . PMC 5411294 . PMID 27980293 .
- ^ Gutiérrez-Vázquez C, Quintana FJ (январь 2018 г.). «Регуляция иммунного ответа рецептором арилглетодородного рецептора» . Иммунитет . 48 (1): 19–33. doi : 10.1016/j.immuni.2017.12.012 . PMC 5777317 . PMID 29343438 .
- ^ Бурбах К.М., Польша А., Брэдфилд Калифорния (сентябрь 1992 г.). «Клонирование кДНК AH-рецептора выявляет отличительный фактор транскрипции, активируемый лигандом» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (17): 8185–8189. Bibcode : 1992pnas ... 89.8185b . doi : 10.1073/pnas.89.17.8185 . PMC 49882 . PMID 1325649 .
- ^ Фукунага Б.Н., Пробст М.Р., Рейс-Порсаш С., Ханкинсон О (декабрь 1995 г.). «Идентификация функциональных доменов арилглетодородного рецептора» . Журнал биологической химии . 270 (49): 29270–29278. doi : 10.1074/jbc.270.49.29270 . PMID 7493958 .
- ^ Джонс С. (2004). «Обзор основных белков спиральной спирали-спирали» . Биология генома . 5 (6): 226. DOI : 10.1186/GB-2004-5-6-226 . PMC 463060 . PMID 15186484 .
- ^ Ema M, Sogawa K, Watanabe N, Chujoh Y, Matsushita N, Gotoh O, et al. (Апрель 1992). «Клонирование кДНК и структура предполагаемого мышиного рецептора AH». Биохимическая и биофизическая исследовательская коммуникация . 184 (1): 246–253. doi : 10.1016/0006-291x (92) 91185-S . PMID 1314586 .
- ^ Jump up to: а беременный Coumailleau P, Poellinger L, Gustafsson JA, Whitelaw ML (октябрь 1995 г.). «Определение минимального домена рецептора диоксина, который связан с HSP90 и поддерживает аффинность и специфичность связывания лиганда дикого типа» . Журнал биологической химии . 270 (42): 25291–25300. doi : 10.1074/jbc.270.42.25291 . PMID 7559670 .
- ^ Goyo K, Suzuki A, Del Carpio CA, Izaki K, Kuryama E, Miam Y, et 1. (Март 2007 г.). «Идентификация аминокислотных остатков в рецепторе AH, участвующих в связывании лиганда» Биохимическая и биофизическая исследовательская коммуникация 354 (2): 396–4 Doi : 10.1016/ j.bbrc.2006.12.2 PMID 17227672
- ^ Кумар М.Б., Рамадосс П., Реен Р.К., Ванден Хевел Дж.П., Пердию Г.Х. (ноябрь 2001 г.). «Поддомен Q-богатого домена трансактивационного рецептора человека необходим для диоксин-опосредованной транскрипционной активности» . Журнал биологической химии . 276 (45): 42302–42310. doi : 10.1074/jbc.m104798200 . PMID 11551916 .
- ^ Денисон М.С., Пандини А., Надь С.Р., Болдуин Эп, Бонати Л (сентябрь 2002 г.). «Связывание лиганда и активация рецептора AH» . Химико-биологические взаимодействия (представленная рукопись). 141 (1–2): 3–24. Bibcode : 2002cbi ... 141 .... 3d . doi : 10.1016/s0009-2797 (02) 00063-7 . PMID 12213382 . S2CID 29379967 .
- ^ Jump up to: а беременный в Денисон М.С., Надь С.Р. (2003). «Активация ариловой углеводородной рецептора структурно разнообразными экзогенными и эндогенными химическими веществами». Ежегодный обзор фармакологии и токсикологии . 43 : 309–334. doi : 10.1146/annurev.pharmtox.43.100901.135828 . PMID 12540743 .
- ^ Baker Jr, Sakoff JA, McCluskey A (май 2020). «Ариловой углеводородной рецептор (AHR) как мишень лекарственного средства для рака молочной железы» . Обзоры лекарственных исследований . 40 (3): 972–1001. doi : 10.1002/med.21645 . PMID 31721255 .
- ^ Adachi J, Mori Y, Matsui S, Takigami H, Fujino J, Kitagawa H, et al. (Август 2001 г.). «Индирубин и индиго являются мощными лигандами арилглетодородных рецепторов, присутствующих в человеческой моче» . Журнал биологической химии . 276 (34): 31475–31478. doi : 10.1074/jbc.c100238200 . PMID 11425848 .
- ^ Синальный CJ, Бенд -младший (октябрь 1997 г.). «Арилглетоглеродная рецептор-зависимая индукция CYP1A1 билирубином в клетках гепатомы мыши HEPA 1C1C7». Молекулярная фармакология . 52 (4): 590–599. doi : 10.1124/моль.52.4.590 . PMID 9380021 .
- ^ Seidel SD, Winters GM, Rogers WJ, Ziccardi MH, Li V, Keser B, et al. (2001). «Активация сигнального пути рецептора AH простагландинами». Журнал биохимической и молекулярной токсикологии . 15 (4): 187–196. doi : 10.1002/jbt.16 . PMID 11673847 . S2CID 21953408 .
- ^ McMillan BJ, Bradfield CA (январь 2007 г.). «Рецептор ариловой углеводороды активируется модифицированным липопротеином низкой плотности» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (4): 1412–1417. Bibcode : 2007pnas..104.1412m . doi : 10.1073/pnas.0607296104 . PMC 1783125 . PMID 17227852 .
- ^ Savouret JF, Antenos M, Quesne M, Xu J, Milgrom E, Casper RF (февраль 2001 г.). «7-кетохолестерин является эндогенным модулятором для арилгидроглеродного рецептора» . Журнал биологической химии . 276 (5): 3054–3059. doi : 10.1074/jbc.m005988200 . PMID 11042205 .
- ^ Safe S (ноябрь 2017 г.). «Carbidopa: селективный модулятор рецептора AH (SAHRM)». Биохимический журнал . 474 (22): 3763–3765. doi : 10.1042/bcj20170728 . PMID 29109131 .
- ^ Ченг Й., Джин У, Дэвидсон Л.А., Чаприн Р.С., Джаяраман А., Тамамис П. и др. (Февраль 2017 г.). «Основной момент редактора: 1,4-дигидрокси-2-нафтоевая кислота, полученная из микробных средств и связанные с ними соединения в качестве арилоглеводородочных рецепторных рецепторов/антагонистов: взаимосвязь структурно-активности и моделирование рецепторов» . Токсикологические науки . 155 (2): 458–473. doi : 10.1093/toxsci/kfw230 . PMC 5291215 . PMID 27837168 .
- ^ Инь XF, Чен Дж., Мао В., Ван Й.Х., Чен М.Х. (май 2012 г.). «Селективный модулятор арилглетодородного рецептора 3,3'-дииндолилметан ингибирует рост раковых клеток желудка» . Журнал экспериментальных и клинических исследований рака . 31 (1): 46. DOI : 10.1186/1756-9966-31-46 . PMC 3403951 . PMID 22592002 .
- ^ Wincent E, Shirani H, Bergman J, Rannug U, Janosik T (февраль 2009 г.). «Синтез и биологическая оценка слитых тио- и селенопиранов в качестве новых аналогов индолокарбазола с арилоуглеродным рецептором аффинности». Биоорганическая и лекарственная химия . 17 (4): 1648–1653. doi : 10.1016/j.bmc.2008.12.072 . PMID 19186062 .
- ^ Денисон М.С., Пандини А., Надь С.Р., Болдуин Эп, Бонати Л (сентябрь 2002 г.). «Связывание лиганда и активация рецептора AH» . Химико-биологические взаимодействия . 141 (1–2): 3–24. Bibcode : 2002cbi ... 141 .... 3d . doi : 10.1016/s0009-2797 (02) 00063-7 . PMID 12213382 . S2CID 29379967 .
- ^ Денис М., Cuthill S, Wikström AC, Poellinger L, Gustafsson JA (сентябрь 1988 г.). «Ассоциация рецептора диоксина с МР 90 000 белка теплового шока: структурное родство с рецептором глюкокортикоидов». Биохимическая и биофизическая исследовательская коммуникация . 155 (2): 801–807. doi : 10.1016/s0006-291x (88) 80566-7 . PMID 2844180 .
- ^ Perdew GH (сентябрь 1988 г.). «Ассоциация рецептора AH с белком теплового шока 90 кДа» . Журнал биологической химии . 263 (27): 13802–13805. doi : 10.1016/s0021-9258 (18) 68314-0 . PMID 2843537 .
- ^ Cox MB, Miller CA (март 2004 г.). «Сотрудничество белка теплового шока 90 и P23 в передаче сигналов арилоггкодородного рецептора» . Клеточный стресс и шапероны . 9 (1): 4–20. doi : 10.1379/460.1 . PMC 1065305 . PMID 15270073 .
- ^ Jump up to: а беременный Kazlauskas A, Poellinger L, Pongratz I (май 1999). «Свидетельство о том, что ко-чаперона P23 регулирует реагирование лиганда рецептора диоксина (арилоггдымодородочный)» . Журнал биологической химии . 274 (19): 13519–13524. doi : 10.1074/jbc.274.19.13519 . PMID 10224120 .
- ^ Kazlauskas A, Sundström S, Poellinger L, Pongratz I (апрель 2001 г.). «Комплекс шаперона HSP90 регулирует внутриклеточную локализацию рецептора диоксина» . Молекулярная и клеточная биология . 21 (7): 2594–2607. doi : 10.1128/mcb.21.7.2594-2607.2001 . PMC 86890 . PMID 11259606 .
- ^ Jump up to: а беременный Shetty PV, Bhagwat By, Chan WK (март 2003 г.). «P23 усиливает образование комплекса арилглетодородного рецептора-ДНК». Биохимическая фармакология . 65 (6): 941–948. doi : 10.1016/s0006-2952 (02) 01650-7 . PMID 12623125 .
- ^ Мейер Б.К., Молитва Грант М.Г., Ванден Хевел Дж.П., Пердею Г.Х. (февраль 1998 г.). «Белок 2 вируса гепатита В.-ассоциированность 2 является субъединицей неволененного ядра арилглетогдородного рецептора и проявляет активность энхансеры транскрипции» . Молекулярная и клеточная биология . 18 (2): 978–988. doi : 10.1128/mcb.18.2.978 . PMC 108810 . PMID 9447995 .
- ^ MA Q, Whitlock JP (апрель 1997 г.). «Новый цитоплазматический белок, который взаимодействует с рецептором AH, содержит тетратропептидные повторные мотивы и расширяет транскрипционный ответ на 2,3,7,8,8-тетрахлордибензо-p-диоксин» . Журнал биологической химии . 272 (14): 8878–8884. doi : 10.1074/jbc.272.14.8878 . PMID 9083006 .
- ^ Jump up to: а беременный Zhou Q, Lavorgna A, Bowman M, Hiscott J, Harhaj EW (июнь 2015 г.). «Интерактивное белок арилоглеводородного рецептора нацелен на IRF7 для подавления антивирусной передачи сигналов и индукции интерферона типа I» . Журнал биологической химии . 290 (23): 14729–14739. doi : 10.1074/jbc.m114.633065 . PMC 4505538 . PMID 25911105 .
- ^ Карвер Л.А., Брэдфилд Калифорния (апрель 1997 г.). «Лиганд-зависимый взаимодействие арилоггдыглеродного рецептора с новым иммунофилиновым гомологом in vivo» . Журнал биологической химии . 272 (17): 11452–11456. doi : 10.1074/jbc.272.17.11452 . PMID 9111057 .
- ^ Carver LA, Jackiw V, Bradfield CA (декабрь 1994 г.). «Белок теплового шока 90 кДа необходим для передачи сигналов рецептора AH в системе экспрессии дрожжей» . Журнал биологической химии . 269 (48): 30109–30112. doi : 10.1016/s0021-9258 (18) 43782-9 . PMID 7982913 .
- ^ Pongratz I, Mason GG, Poellinger L (июль 1992 г.). «Двойная роль белка теплового шока 90 кДа HSP90 в модулировании функциональной активности рецептора диоксина. Свидетельство о том, что рецептор диоксина, функционально относящийся к подклассу ядерных рецепторов, которые требуют HSP90 как для активности связывания лиганда, так и для репрессии внутренней связывающей активности ДНК», которые требуют HSP90 как для лиганда, так и для репрессии внутренней ДНК-активности », которые требуют HSP90 как для активности связывания лигандов, так и для репрессии внутренней ДНК-активности». Полем Журнал биологической химии . 267 (19): 13728–13734. doi : 10.1016/s0021-9258 (18) 42274-0 . PMID 1320028 .
- ^ Jump up to: а беременный в Whitelaw M, Pongratz I, Wilhelmsson A, Gustafsson JA, Poellinger L (апрель 1993 г.). «Лиганд-зависимый рекрутинг корректатора ARNT определяет распознавание ДНК рецептором диоксина» . Молекулярная и клеточная биология . 13 (4): 2504–2514. doi : 10.1128/mcb.13.4.2504 . PMC 359572 . PMID 8384309 .
- ^ Карвер Л.А., Лапрес Дж.Дж., Джайн С., Данхэм Е.Е., Брэдфилд Калифорния (декабрь 1998 г.). «Характеристика белка, ассоциированного с рецептором AH, ARA9» . Журнал биологической химии . 273 (50): 33580–33587. doi : 10.1074/jbc.273.50.33580 . PMID 9837941 .
- ^ Петрулис -младший, Ордо Нг, Пердию Г.Х. (декабрь 2000 г.). «Субклеточная локализация рецептора ариловой углеводороды модулируется вирусом H-ассоциированного вируса гепатита B-иммунофилина . Журнал биологической химии . 275 (48): 37448–37453. doi : 10.1074/jbc.m006873200 . PMID 10986286 .
- ^ Петрулис Дж.Р., Куснади А., Рамадосс П., Холлингсхед Б., Пердею Г.Х. (январь 2003 г.). «HSP90 Co-Chaperone XAP2 изменяет импортирование бета-распознавания двухпартного сигнала локализации ядерной локализации рецептора AH и подавляет транскрипционную активность» . Журнал биологической химии . 278 (4): 2677–2685. doi : 10.1074/jbc.m209331200 . PMID 12431985 .
- ^ Jump up to: а беременный Mimura J, Fujii-Kuriyama Y (февраль 2003 г.). «Функциональная роль AHR в экспрессии токсических эффектов с помощью TCDD». Biochimica et Biophysica Acta (BBA) - Общие субъекты . Клеточная биология регуляции цитохрома P450. 1619 (3): 263–268. doi : 10.1016/s0304-4165 (02) 00485-3 . PMID 12573486 .
- ^ Пуга А., Томлинсон К.Р., Ся Ю (январь 2005 г.). «Ах рецептор сигнализирует о перекрестном разговоре с множественными путями развития». Биохимическая фармакология . 69 (2): 199–207. doi : 10.1016/j.bcp.2004.06.043 . PMID 15627472 .
- ^ Puga A, Ma C, Marlowe JL (февраль 2009 г.). «Поперечные разборы рецептора ариловой углеводороды с множественными путями трансдукции сигнальной трансдукции» . Биохимическая фармакология . 77 (4): 713–722. doi : 10.1016/j.bcp.2008.08.031 . PMC 2657192 . PMID 18817753 .
- ^ Икута Т., Эгучи Х., Тачибана Т., Йонда Ю., Каваджири К (январь 1998 г.). «Ядерная локализация и экспортные сигналы арилоглеродообразного рецептора человека» человека » . Журнал биологической химии . 273 (5): 2895–2904. doi : 10.1074/jbc.273.5.2895 . PMID 9446600 .
- ^ PollaNz RS, Barbour ER (август 2000 г.). «Анализ сложной взаимосвязи между ядерной экспортом и регуляцией гена опосредованной арилглетодородным рецептором» . Молекулярная и клеточная биология . 20 (16): 6095–6104. doi : 10.1128/mcb.20.16.6095-6104.2000 . PMC 86085 . PMID 10913191 .
- ^ Хоффман Е.К., Рейес Х., Чу Ф.Ф., Сандер Ф., Конли Л.Х., Брукс Б.А. и др. (Май 1991). «Клонирование фактора, необходимого для активности рецептора AH (диоксин)». Наука . 252 (5008): 954–958. Bibcode : 1991sci ... 252..954H . doi : 10.1126/science.1852076 . PMID 1852076 .
- ^ Jump up to: а беременный Пробст М.Р., Рейс-Порсаш С., Агбунаг Р.В., Онг М.С., Ханкинсон О (сентябрь 1993 г.). «Роль белка ядерного транслокатора арилглетодородного рецептора в рецепторе арилглетодородного (диоксин)». Молекулярная фармакология . 44 (3): 511–518. PMID 8396713 .
- ^ Рейес Х., Рейс-Порсаш С., Ханкинсон О (май 1992). «Идентификация белка ядерного транслокатора рецептора AH (ARNT) в качестве компонента формы связывания ДНК рецептора AH». Наука . 256 (5060): 1193–1195. Bibcode : 1992sci ... 256.1193r . doi : 10.1126/science.256.5060.1193 . PMID 1317062 . S2CID 34075046 .
- ^ Dolwick KM, Swanson HI, Bradfield CA (сентябрь 1993 г.). «Анализ in vitro доменов рецептора AH, участвующих в распознавании ДНК-активированного лиганда» » . Труды Национальной академии наук Соединенных Штатов Америки . 90 (18): 8566–8570. Bibcode : 1993pnas ... 90.8566d . doi : 10.1073/pnas.90.18.8566 . PMC 47398 . PMID 8397410 .
- ^ Shen ES, Whitlock JP (апрель 1992 г.). «Взаимодействие белка-ДНК в энхансере, чувствительном к диоксинам. Мутационный анализ ДНК-связывающего сайта для рецептора AH с лигандом» . Журнал биологической химии . 267 (10): 6815–6819. doi : 10.1016/s0021-9258 (19) 50499-9 . PMID 1313023 .
- ^ Lusska A, Shen E, Whitlock JP (март 1993 г.). «Взаимодействие белка-ДНК в энхансере, чувствительном к диоксинам. Анализ шести добросовестных ДНК-связывающих сайтов для рецептора AH с лигитом» . Журнал биологической химии . 268 (9): 6575–6580. doi : 10.1016/s0021-9258 (18) 53289-0 . PMID 8384216 .
- ^ Яо Эф, Денисон М.С. (июнь 1992 г.). «Детерминанты последовательности ДНК для связывания трансформированного рецептора AH с энхансером, чувствительным к диоксинам». Биохимия . 31 (21): 5060–5067. doi : 10.1021/bi00136a019 . PMID 1318077 .
- ^ Wharton KA, Franks RG, Kasai Y, Crews St (декабрь 1994 г.). «Контроль транскрипции средней линии CNS с помощью асимметричных электрон-похожих элементов: сходство с ксенобиотической регуляцией» . Разработка . 120 (12): 3563–3569. doi : 10.1242/dev.120.12.3563 . PMID 7821222 .
- ^ Bacsi SG, Reisz-Porszasz S, Hankinson O (март 1995 г.). «Ориентация рецепторного комплекса гетеродимерного рецептора арилоуглеродов (диоксин) на его асимметричной последовательности распознавания ДНК». Молекулярная фармакология . 47 (3): 432–438. PMID 7700240 .
- ^ Swanson HI, Chan WK, Bradfield CA (ноябрь 1995 г.). «Специфичность связывания ДНК и правила спаривания белков рецептора AH, ARNT и SIM» . Журнал биологической химии . 270 (44): 26292–26302. doi : 10.1074/jbc.270.44.26292 . PMID 7592839 .
- ^ Boutros PC, Moffat ID, Franc Ma, Tijet N, Tuomisto J, Pohjanvirta R, et al. (Август 2004 г.). «Аккумулятор гена AHRE-II, реагирующая на диоксин: идентификация с помощью филогенетической отпечатки». Биохимическая и биофизическая исследовательская коммуникация . 321 (3): 707–715. doi : 10.1016/j.bbrc.2004.06.177 . PMID 15358164 .
- ^ Согава К., Нумаяма-Цурута К., Такахаши Т., Мацушита Н., Миура С., Никава Дж. И др. (Июнь 2004 г.). «Новый индукционный механизм крыс CYP1A2, родившийся гетеродимером AH рецептора-артизатора» Биохимическая и биофизическая исследовательская коммуникация 318 (3): 746–7 Doi : 10.1016/ j.bbrc.2004.04.0 15144902PMID
- ^ Hahn Me, Karchner SI, Evans BR, Franks DG, Merson RR, Laperitis JM (сентябрь 2006 г.). «Неожиданное разнообразие арильных углеводородных рецепторов у не млекопитающих позвоночных: понимание сравнительной геномики». Журнал экспериментальной зоологии, часть A: сравнительная экспериментальная биология . 305 (9): 693–706. Bibcode : 2006jeza..305..693h . doi : 10.1002/jez.a.323 . PMID 16902966 .
- ^ Дункан Д.М., Берджесс Е.А., Дункан I (май 1998). «Контроль дистальной антенной идентичности и развития лапки у дрозофилы с помощью беззаношной аристапии, гомолога рецептора диоксина млекопитающих» . Гены и развитие . 12 (9): 1290–1303. doi : 10.1101/gad.12.9.1290 . PMC 316766 . PMID 9573046 .
- ^ Emmons RB, Duncan D, Estes PA, Kiefel P, Mosher JT, Sonnenfeld M, et al. (Сентябрь 1999). «Белки белки без шиковины и танго BHLH-PAS взаимодействуют, чтобы контролировать развитие антенна и лапа у дрозофилы» . Разработка . 126 (17): 3937–3945. doi : 10.1242/dev.126.17.3937 . PMID 10433921 .
- ^ Jump up to: а беременный Tijet N, Boutros PC, Moffat ID, Okey AB, Tuomisto J, Pohjanvirta R (январь 2006 г.). «Арилглеводородный рецептор регулирует различные диоксин-зависимые и диоксин-независимые генные батареи». Молекулярная фармакология . 69 (1): 140–153. doi : 10.1124/моль.105.018705 . PMID 16214954 . S2CID 1913812 .
- ^ Gasiewicz TA, Singh KP, Casado FL (март 2010 г.). «Рецептор арилглетодорода играет важную роль в регуляции гематопоэза: последствия для бензола-индуцированной гематопоэтической токсичности» . Химико-биологические взаимодействия . 184 (1–2): 246–251. Bibcode : 2010cbi ... 184..246g . doi : 10.1016/j.cbi.2009.10.019 . PMC 2846208 . PMID 19896476 .
- ^ KISS EA, Vonarbourg C, Kopfmann S, Hobeika E, Finke D, Esser C, et al. (Декабрь 2011 г.). «Природные лиганды арилоглеводородных рецепторов контролируют органогенез кишечных лимфоидных фолликулов». Наука . 334 (6062): 1561–1565. Bibcode : 2011sci ... 334.1561K . doi : 10.1126/science.1214914 . PMID 22033518 . S2CID 206537957 .
- ^ Jump up to: а беременный Li Y, Innocentin S, Withers DR, Roberts NA, Gallagher AR, Grigorieva EF, et al. (Октябрь 2011). «Экзогенные стимулы поддерживают интраэпителиальные лимфоциты посредством активации арилглетодородного рецептора» . Клетка . 147 (3): 629–640. doi : 10.1016/j.cell.2011.09.025 . PMID 21999944 . S2CID 16090460 .
- ^ Кинтана Ф.Дж., Бассо А.С., Иглесиас А.Х., Корн Т., Фарес М.Ф., Беттелли Е. и др. (Май 2008 г.). «Контроль T (reg) и T (H) 17 дифференцировки клеток арилоглетодородным рецептором». Природа . 453 (7191): 65–71. doi : 10.1038/nature06880 . PMID 18362915 . S2CID 4384276 .
- ^ Акахоши Э., Йошимура С., Ишихара-Сугано М (сентябрь 2006 г.). «Чрезмерная экспрессия AHR (арилоггронодородный рецептор) индуцирует нейронную дифференцировку клеток нейро2A: исследование нейротоксикологии» . Здоровье окружающей среды . 5 (1): 24. Bibcode : 2006envhe ... 5 ... 24a . doi : 10.1186/1476-069x-5-24 . PMC 1570454 . PMID 16956419 .
- ^ Walisser JA, Glover E, Pande K, Liss AL, Bradfield CA (декабрь 2005 г.). «Арилглетогбородное рецептор-зависимое развитие печени и гепатотоксичность опосредованы различными типами клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (49): 17858–17863. Bibcode : 2005pnas..10217858W . doi : 10.1073/pnas.0504757102 . PMC 1308889 . PMID 16301529 .
- ^ Boitano AE, Wang J, Romeo R, Bouchez LC, Parker AE, Sutton SE, et al. (Сентябрь 2010). «Антагонисты арилглеводородов рецепторов способствуют расширению гематопоэтических стволовых клеток человека» . Наука . 329 (5997): 1345–1348. Bibcode : 2010sci ... 329.1345b . doi : 10.1126/science.1191536 . PMC 3033342 . PMID 20688981 .
- ^ Линдси С., Папоутскис и др. (Февраль 2011 г.). «Фактор транскрипции арилглетодородного рецептора (AHR) регулирует мегакариоцитарную полиплоидризацию» . Британский журнал гематологии . 152 (4): 469–484. doi : 10.1111/j.1365-2141.2010.08548.x . PMC 3408620 . PMID 21226706 .
- ^ Merchak A, Gaultier A (декабрь 2020 г.). «Микробные метаболиты и иммунная регуляция: новые мишени при серьезных депрессивных расстройствах» . Мозг, поведение и иммунитет - здоровье . 9 : 100169. DOI : 10.1016/j.bbih.2020.100169 . PMC 8474524 . PMID 34589904 .
- ^ Израиль Д.П., Уитлок Дж.П. (сентябрь 1983 г.). «Индукция мРНК, специфичной для цитохрома P1-450 в клетках гепатомы дикого типа и варианты мыши» . Журнал биологической химии . 258 (17): 10390–10394. doi : 10.1016/s0021-9258 (17) 44469-3 . PMID 6885786 .
- ^ Израиль Ди, Уитлок Дж.П. (май 1984). «Регуляция транскрипции гена цитохрома P1-450 на 2,3,7, 8-тетрахлордибензо-п-диоксин в клетках гепатомы дикого типа и варианты мыши» . Журнал биологической химии . 259 (9): 5400–5402. doi : 10.1016/s0021-9258 (18) 91022-7 . PMID 6715350 .
- ^ KO HP, Okino St, MA Q, Whitlock JP (январь 1996 г.). «Транскрипция CYP1A1, индуцированная диоксином, in vivo: рецептор ароматического углеводорода опосредует трансактивацию, коммуникацию-энхансер-промотор и изменения в структуре хроматина» . Молекулярная и клеточная биология . 16 (1): 430–436. doi : 10.1128/mcb.16.1.430 . PMC 231019 . PMID 8524325 .
- ^ Nebert DW, Roe AL, Dieter MZ, Solis WA, Yang Y, Dalton TP (январь 2000 г.). «Роль ароматического углеводородного рецептора и генной батареи [AH] в реакции окислительного стресса, контроля клеточного цикла и апоптоза». Биохимическая фармакология . 59 (1): 65–85. doi : 10.1016/s0006-2952 (99) 00310-x . PMID 10605936 .
- ^ Lensu S, Tuomisto JT, Tuomisto J, Viluksela M, Niittynen M, Pohjanvirta R (июнь 2011 г.). «Непосредственная и высокочувствительная реакция на отвращение на новый продукт питания, связанный с стимуляцией рецептора AH». Токсикологические письма . 203 (3): 252-257. Doi : 10.1016/j.toxlet.2011.03.025 . PMID 21458548 .
- ^ Monteleone I, Rizzo A, Sarra M, Sica G, Sileri P, Biancone L, et al. (Июль 2011). «Индуцированные арилоглеводородочными рецепторами сигналы повышают регулирующие продукты IL-22 и ингибируют воспаление в желудочно-кишечном тракте» . Гастроэнтерология . 141 (1): 237–48, 248.e1. doi : 10.1053/j.gastro.2011.04.007 . PMID 21600206 .
- ^ Wei P, Hu GH, Kang HY, Yao HB, Kou W, Liu H, et al. (Май 2014). «Лиганд рецептора ариловой углеводороды действует на дендритные клетки и Т -клетки для подавления ответа Th17 у пациентов с аллергическим ринитом» . Лабораторное расследование; Журнал технических методов и патологии . 94 (5): 528–535. doi : 10.1038/labinvest.2014.8 . PMID 24514067 .
- ^ Memari B, Bouttier M, Dimitrov V, Ouellette M, Behr MA, Fritz JH, et al. (Ноябрь 2015). «Взаимодействие ариловой углеводородной рецептора у макрофагов, инфицированных микобактерием туберкулеза, обладает плейотропной влиянием на врожденную иммунную передачу» . Журнал иммунологии . 195 (9): 4479–4491. doi : 10.4049/jimmunol.1501141 . PMID 26416282 .
- ^ Harrigan JA, Vezina CM, McGarrigle BP, Ersing N, Box HC, MacCubbin AE, et al. (Февраль 2004 г.). «Образование аддукта ДНК в печени крысы с точностью и срезы легких, подвергшихся воздействию бензо [а] пирена» . Токсикологические науки . 77 (2): 307–314. doi : 10.1093/toxsci/kfh030 . PMID 14691214 .
- ^ Lindén J, Lensu S, Tuomisto J, Pohjanvirta R (октябрь 2010). «Диоксины, рецептор арила углеводородов и центральная регуляция энергетического баланса». Границы в нейроэндокринологии . 31 (4): 452–478. doi : 10.1016/j.yfrne.2010.07.002 . PMID 20624415 . S2CID 34036181 .
- ^ Martinez JM, Afshari CA, Bushel PR, Masuda A, Takahashi T, Walker NJ (октябрь 2002 г.). «Дифференциальные токсикогеномные реакции на 2,3,7,8-тетрахлордибензо-п-диоксин в злокачественных и незлокачественных эпителиальных клетках дыхательных путей человека» . Токсикологические науки . 69 (2): 409–423. doi : 10.1093/toxsci/69.2.409 . PMID 12377990 .
- ^ Vezina CM, Walker NJ, Olson Jr (ноябрь 2004 г.). «Подхроническое воздействие TCDD, PECDF, PCB126 и PCB153: влияние на экспрессию генов печени» . Перспективы здоровья окружающей среды . 112 (16): 1636–1644. doi : 10.1289/ehp.7253 . PMC 1247661 . PMID 15598615 .
- ^ Овандо Б.Дж., Везина С.М., МакГарригл Б.П., Олсон -младший (декабрь 2006 г.). «Понижающая регуляция гена печени после острого и субхронического воздействия 2,3,7,8-тетрахлордибензо-п-диоксин» . Токсикологические науки . 94 (2): 428–438. doi : 10.1093/toxsci/kfl111 . PMID 16984957 .
- ^ Hogenesch JB, Chan WK, Jackiw VH, Brown RC, Gu YZ, Pray-Grant M, et al. (Март 1997 г.). «Характеристика подмножества суперсемейства с базовой спиралью-петли-спиралью, которая взаимодействует с компонентами пути сигнализации диоксина» . Журнал биологической химии . 272 (13): 8581–8593. doi : 10.1074/jbc.272.13.8581 . PMID 9079689 .
- ^ Tian Y, Ke S, Chen M, Sheng T (ноябрь 2003 г.). «Взаимодействие между арилоггдыгдородным рецептором и P-TEFB. Последовательный рекрутирование факторов транскрипции и дифференциальным фосфорилированием С-концевого домена РНК-полимеразы II при промоторе CYP1A1» . Журнал биологической химии . 278 (45): 44041–44048. doi : 10.1074/jbc.m306443200 . PMID 12917420 .
- ^ Wormke M, Stoner M, Saville B, Walker K, Abdelrahim M, Burghardt R, et al. (Март 2003 г.). «Рецептор арилглетодорода опосредует деградацию альфа рецептора эстрогена посредством активации протеасомов» . Молекулярная и клеточная биология . 23 (6): 1843–1855. doi : 10.1128/mcb.23.6.1843-1855.2003 . PMC 149455 . PMID 12612060 .
- ^ Klinge CM, Kaur K, Swanson HI (январь 2000 г.). «Рецептор арилглетодорода взаимодействует с рецепторами эстрогена альфа и рецепторами сироты Coup-TFI и Erlpha1». Архивы биохимии и биофизики . 373 (1): 163–174. doi : 10.1006/abbi.1999.1552 . PMID 10620335 .
- ^ Beischlag TV, Wang S, Rose DW, Torchia J, Reisz-Porszasz S, Muhammad K, et al. (Июнь 2002 г.). «Рекрутирование семейства транскрипционных коактиваторов NCOA/SRC-1/P160 с помощью арилоггдыглетодородного рецептора/арилоггдыкоглеродистого рецептора ядерного транслокаторного комплекса» . Молекулярная и клеточная биология . 22 (12): 4319–4333. doi : 10.1128/mcb.22.12.4319-4333.2002 . PMC 133867 . PMID 12024042 .
- ^ Antenos M, Casper RF, Brown TJ (ноябрь 2002 г.). «Взаимодействие с NEDD8, убиквитиноподобным белком, усиливает транскрипционную активность рецептора ариловой углеводороды» . Журнал биологической химии . 277 (46): 44028–44034. doi : 10.1074/jbc.m202413200 . PMID 12215427 .
- ^ Kumar MB, Tarpey RW, Perdew GH (август 1999 г.). «Дифференциальный рекрутинг коактиватора RIP140 с помощью рецепторов AH и эстрогена. Отсутствие роли для мотивов LXXLL» . Журнал биологической химии . 274 (32): 22155–22164. doi : 10.1074/jbc.274.32.22155 . PMID 10428779 .
- ^ Kim DW, Gazourian L, Quadri SA, Romieu-Mourez R, Sherr DH, Sonenshein GE (ноябрь 2000 г.). «Субъединица Rela-kappab и арилоггдыглеточный рецептор (AHR) сотрудничают для трансактивации промотора C-Myc в клетках молочной железы» . Онкоген . 19 (48): 5498–5506. doi : 10.1038/sj.onc.1203945 . PMID 11114727 .
- ^ Ruby CE, Leid M, Kerkvliet Ni (сентябрь 2002 г.). «2,3,7,8-тетрахлордибензо-P-диоксин подавляет фактор некроза опухоли-альфа и анти-CD40, индуцированную активацией NF-KAPPAB/REL в дендритных клетках: активация гомодимера P50 не влияет». Молекулярная фармакология . 62 (3): 722–728. doi : 10.1124/моль.62.3.722 . PMID 12181450 . S2CID 2376236 .
- ^ Vogel CF, Sciullo E, Li W, Wong P, Lazennec G, Matsumura F (декабрь 2007 г.). «RELB, новый партнер транскрипции, опосредованной арил углеводородов» . Молекулярная эндокринология . 21 (12): 2941–2955. doi : 10.1210/me.2007-0211 . PMC 2346533 . PMID 17823304 .
- ^ GE NL, Elferink CJ (август 1998 г.). «Прямое взаимодействие между рецептором ариловой углеводороды и белком ретинобластомы. Связывание передачи сигналов диоксина с клеточным циклом» . Журнал биологической химии . 273 (35): 22708–22713. doi : 10.1074/jbc.273.35.22708 . PMID 9712901 .
Внешние ссылки
[ редактировать ]- Арил+углеводородный+рецептор в Национальной библиотеке медицинской библиотеки Медицинской библиотеки США (Mesh)
- Human AHR Местоположение генома AHR и страница деталей гена в браузере генома UCSC .
- страница . Расположение генома ARNT и ARNT генов UCSC Genome в браузере