Связанный с RAR рецептор-сирот альфа
| Рора | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RORA , NR1F1, ROR1, ROR2, ROR3, RZR-ALPHA, RZRA, RAR, связанный с сиротой рецептор A, Iddeca | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 600825 ; MGI : 104661 ; Гомологен : 56594 ; Genecards : Rora ; OMA : RORA - Ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




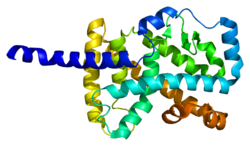
Связанный с RAR альфа-рецептор-сирот ( RORα ), также известный как NR1F1 (подсемейство 1-я ядерного рецептора 1, группа F, член 1), является ядерным рецептором , который у людей кодируется RORA геном . [ 5 ] RORα участвует в регуляции транскрипции некоторых генов, участвующих в циркадном ритме . [ 6 ] У мышей RORα необходим для развития мозжечка [ 7 ] [ 8 ] посредством прямой регуляции генов, экспрессируемых в клетках Пуркинье. [ 9 ] Он также играет важную роль в развитии врожденных лимфоидных клеток типа 2 (ILC2), а мутантные животные дефицит ILC2. [ 10 ] [ 11 ] Кроме того, хотя клетки ILC3 и Th17 от мышей с дефицитом RORα у мышей, дефицитных, дефектны для цитокинов . выработки [ 12 ]
Открытие
[ редактировать ]Первые трехместные изоформы RORα были первоначально клонированы и охарактеризованы как ядерные рецепторы в 1994 году Giguère и коллегами, когда их структура и функция были впервые изучены. [ 13 ]
В начале 2000 -х годов различные исследования показали, что RORα демонстрирует ритмические паттерны экспрессии в циркадном цикле в печени , почках , сетчатке и легких . [ 14 ] Интересно, что примерно в это же время было обнаружено, что численность RORα является циркадным в супрахиазматическом ядре млекопитающих . [ 15 ] RORα необходим для нормальных циркадных ритмов у мышей , [ 16 ] демонстрируя его важность в хронобиологии .
Структура
[ редактировать ]Белок, кодируемый этим геном, является членом подсемейства NR1 рецепторов ядерных гормонов. [ 16 ] У людей было идентифицировано 4 изоформы RORα, которые генерируются посредством альтернативного сплайсинга и использования промотора и демонстрируют дифференциальную ткань-специфическую экспрессию. Структура белка RORα состоит из четырех канонических функциональных групп: N -концевого (A/B) домена, ДНК -связывающего домена , содержащего два цинковых пальца , домен шарнира и C -концевой лиганд -связывающий домен. В семье ROR ДНК-связывающий домен высоко консервативен, а лиганд-связывающий домен только умеренно консервативный. [ 14 ] Различные изоформы RORα имеют различную специфичность связывания и сильные стороны транскрипционной активности. [ 5 ]
Регуляция циркадного ритма
[ редактировать ]Основные млекопитающих циркадные часы - это отрицательная петля обратной связи , которая состоит из PER1/PER2 , CRY1/CRY2 , BMAL1 и часов . [ 15 ] Эта обратная связь стабилизируется через другую петлю, включающую транскрипционную регуляцию BMAL1 . [ 17 ] Трансактивация BMAL1 промоторе регулируется через восходящий элемент ответа ROR/Rev-ERB (RRE) в BMAL1 , с которым связывают RORα и Rev-ERBα . [ 17 ] Эта стабилизирующая регуляторная петля индуцируется гетеродимером BMAL1/часов , который индуцирует транскрипцию RORα и Rev-ERBα . [ 15 ] RORα, который активирует транскрипцию BMAL1 , и Rev-ERBα, который подавляет транскрипцию BMAL1 , конкурирует, чтобы связываться с RRE. [ 17 ] эта петля обратной связи, регулирующая экспрессию BMAL1 Считается, что , стабилизирует механизм основных часов, помогая сформировать его от изменений в окружающей среде . [ 17 ]
Механизм
[ редактировать ]Конкретная связь с элементами ROR (RORE) в регуляторных областях необходима для функции RORα в качестве активатора транскрипции. [ 18 ] RORα достигает этого путем специфического связывания с консенсусным мотивом Core в RORE, RGGTCA. Это взаимодействие возможно благодаря ассоциации первого цинкового пальца RORα с основным мотивом в основной канавке, P-боксе и ассоциации его C-концевого расширения с AT-богатой областью в 5-футовой области RORE. [ 16 ]
Гомология
[ редактировать ]RORα, RORβ и RORγ являются транскрипционными активаторами, распознавающими элементы ROR-ответа. [ 19 ] ROR-альфа экспрессируется в различных типах клеток и участвует в регуляции нескольких аспектов развития, воспалительных реакций и развития лимфоцитов . [ 20 ] Изоформы RORα (RORα1 через RORα3) возникают посредством альтернативной обработки РНК, причем RORα2 и RORα3 разделяют аминоконцевую область, отличную от RORα1. [ 5 ] В отличие от RORα, RORβ экспрессируется в тканях центральной нервной системы (CNS), участвующих в обработке сенсорной информации и в генерировании циркадных ритмов, в то время как RORγ является критическим в лимфатических узлов организме и тимопоэизисе . [ 20 ]
ДНК-связывающие домены рецептора сирот DHR3 у дрозофилы показывают особенно тесную гомологию в амино и карбокси , прилегающих к второй области цинкового пальца в RORα, что позволяет предположить, что эта группа остатков важна для функциональности белков. [ 5 ]
PDP1 и VRI в Drosophila регулируют циркадный ритм, конкурируя за тот же сайт связывания, VP-короб, аналогично тому, как ROR и Rev-Erb конкурентно связываются с RRE. [ 17 ] PDP1 и VRI представляют собой петлю обратной связи и являются функциональными гомологами ROR и Rev-ERB у млекопитающих. [ 17 ]
Прямые ортологи этого гена были идентифицированы у мышей и людей.
Цитохром C -псевдоген HC2 и RORα разделяют перекрывающуюся геномную организацию с псевдогеном HC2, расположенным в пределах транскрипции RORα2. Нуклеотидные и выведенные аминокислотные последовательности псевдогена цитохрома-С-обработки находятся на цепи смысла , в то время как последователи амино-концевого экзона RORα2 находятся на антисмысловой цепи. [ 5 ]
Взаимодействия
[ редактировать ]- ДНК: RORα связывается с P-box RORE. [ 16 ]
- Коактиваторы :
- SRC-1 , CBP , P300 , Trip-L, Trip-230 , промежуточный протеин-1 транскрипции ( TIF-1 ), пролифератор-связывающий пролифератор белок (PBP) и GRIP-1 физически взаимодействуют с RORα. [ 14 ]
- Мотив LXXLL: ROR взаимодействует с SRC-1, GRIP-L, CBP и P300 через мотивы LXXLL (L = Leucine, x = любая аминокислота) на этих белках. [ 14 ]
- SRC-1 , CBP , P300 , Trip-L, Trip-230 , промежуточный протеин-1 транскрипции ( TIF-1 ), пролифератор-связывающий пролифератор белок (PBP) и GRIP-1 физически взаимодействуют с RORα. [ 14 ]
- Убиквитинирование : RORα нацелен на протеасому путем убиквитинирования. Ко -э-берпрессор , безволосый, стабилизирует RORα, защищая его от этого процесса, который также подавляет RORα. [ 21 ]
- Sumoylation : UBE21/UBC9: Убиквитин-конъюгирующий фермент I взаимодействует с RORS, но его эффект еще не известен. [ 16 ]
- Фосфорилирование :
- Фосфорилирование RORα1, которое ингибирует его транскрипционную активность, индуцируется протеинкиназой C. [ 14 ]
- ERK2 : внеклеточный сигнал, регулируемая киназа-2, также фосфорилирует RORα. [ 22 ]
- ATXN1 : ATXN1 и RORα образуют часть белкового комплекса в клетках Purkinje . [ 16 ]
- FOXP3 : FOXP3 напрямую подавляет транскрипционную активность RORS. [ 16 ]
- NME1 : ROR, как было показано, специфически взаимодействует с NME1. [ 23 ]
- NM23-2 : NM23-2 является нуклеозидной дифхосфаткиназой, участвующей в органогенезе и дифференцировке. [ 6 ]
- NM23-1 : NM23-1 является продуктом гена- супрессора-супрессора опухоли . [ 6 ]
Как цель наркотиков
[ редактировать ]Поскольку RORα и Rev-ERBα являются ядерными рецепторами, которые имеют одни и те же гены-мишени и участвуют в процессах, которые регулируют метаболизм , развитие , иммунитет и циркадный ритм, они демонстрируют потенциал в качестве лекарственных целей . Синтетические лиганды имеют различные потенциальные терапевтические применения и могут использоваться для лечения таких заболеваний, как диабет , атеросклероз , аутоиммунитет и рак . Было обнаружено, что T0901317 и SR1001, два синтетических лиганда, являются RORα и RORγ обратными агонистами , которые подавляют репортерную активность и, как было показано, задерживают начало и клиническую тяжесть рассеянного склероза и других Th17 клеточных клеточных, опосредованных клетками . SR1078 был обнаружен в качестве агониста RORα и RORγ, который увеличивает экспрессию G6PC и FGF21, что дает терапевтический потенциал для лечения ожирения и диабета, а также рака молочной железы , яичников и простаты . SR3335 также был обнаружен в качестве обратного агониста RORα. [ 13 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000069667 - ENSEMBL , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000032238 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Jump up to: а беременный в дюймовый и Giguère V, Tini M, Flock G, Ong E, Evans RM, Otulakowski G (март 1994 г.). «Изоформные аминоконцевые домены диктуют ДНК-связывающие свойства ROR Alpha, нового семейства ядерных рецепторов-сирот» . Гены и развитие . 8 (5): 538–53. doi : 10.1101/gad.8.5.538 . PMID 7926749 .
- ^ Jump up to: а беременный в «Ген Entrez: Rora RAR-рецептор-сирот» А » .
- ^ Сидман Р.Л., Лейн П.В., Дики М.М. (август 1962 г.). «Шофляер, новая мутация в мыши, затрагивающая мозжечок». Наука . 137 (3530): 610–2. Bibcode : 1962sci ... 137..610S . doi : 10.1126/science.137.3530.610 . PMID 13912552 . S2CID 30733570 .
- ^ Гамильтон Б.А., Франкель В.Н., Керреброк А.В., Хокинс Т.Л., Фицхью В., Кусуми К. и др. (Февраль 1996 г.). «Разрушение рецептора ядерного гормона roralpha ускоростных мышей». Природа . 379 (6567): 736–9. Bibcode : 1996natur.379..736h . doi : 10.1038/379736A0 . PMID 8602221 . S2CID 4318427 .
- ^ Gold DA, Baek SH, Schork NJ, Rose DW, Larsen DD, Sachs BD, et al. (Декабрь 2003 г.). «Roralpha координирует взаимную передачу сигналов в развитии мозжечка с помощью звукового ежи и кальций-зависимых путей» . Нейрон . 40 (6): 1119–31. doi : 10.1016/s0896-6273 (03) 00769-4 . PMC 2717708 . PMID 14687547 .
- ^ Халим Тай, Макларен А., Романиш М.Т., Голд М.Дж., Макнагни К.М., Такеи Ф. (сентябрь 2012 г.). «Альфа-рецептор-рецептор-рецептор, связанный с рецептором-рецептором, требуется для развития природных вспомогательных клеток и аллергического воспаления» . Иммунитет . 37 (3): 463–74. doi : 10.1016/j.immuni.2012.06.012 . PMID 22981535 .
- ^ Gold MJ, Antignano F, Halim Ty, Hirota JA, Blanchet MR, Zaph C, et al. (Апрель 2014). «Группа 2 врожденные лимфоидные клетки облегчают сенсибилизацию к локальному, но не системному, индуцирующему Th2-индикационному воздействию аллергенов». Журнал аллергии и клинической иммунологии . 133 (4): 1142–8. doi : 10.1016/j.jaci.2014.02.033 . PMID 24679471 .
- ^ LO BC, Gold MJ, Hughes MR, Antignano F, Valdez Y, Zaph C, et al. (Сентябрь 2016 г.). «Ядерный рецептор -сирот ROR Alpha и группа 3 врожденные лимфоидные клетки приводят к фиброзу в мышиной модели болезни Крона» . Научная иммунология . 1 (3): EAAF8864. doi : 10.1126/sciimmunol.aaf8864 . PMC 5489332 . PMID 28670633 .
- ^ Jump up to: а беременный Kojetin DJ, Burris TP (март 2014 г.). «Ядерные рецепторы Rev-Erb и ROR в качестве лекарственных целей» . Природные обзоры. Открытие наркотиков . 13 (3): 197–216. doi : 10.1038/nrd4100 . PMC 4865262 . PMID 24577401 .
- ^ Jump up to: а беременный в дюймовый и Jetten AM, Kurebayashi S, Ueda E (2001). «Подсемейство ядерного рецептора ядерного сироты: критические регуляторы множественных биологических процессов» . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии . 69 : 205–47 . doi : 10.1016/s0079-6603 (01) 69048-2 . ISBN 978-0-12-540069-5 Анкет PMID 11550795 .
- ^ Jump up to: а беременный в KO CH, Takahashi JS (октябрь 2006 г.). «Молекулярные компоненты циркадных часов млекопитающих» . Молекулярная генетика человека . 15 Спецификация № 2 (2): R271-7. doi : 10.1093/hmg/ddl207 . PMC 3762864 . PMID 16987893 .
- ^ Jump up to: а беременный в дюймовый и фон Emery P, Reppert SM (август 2004 г.). «Ритмичный рор» . Нейрон . 43 (4): 443–6. doi : 10.1016/j.neuron.2004.08.009 . PMID 15312644 .
- ^ Laitinen S, Staels B (2003). «Потенциальная роль Рор-Альфа в сердечно-сосудистой эндокринологии» . Передача сигналов ядерного рецептора . 1 : E011. doi : 10.1621/nrs.01011 . PMC 1402228 . PMID 16604183 .
- ^ Zhao X, Cho H, Yu RT, Atkins AR, Downes M, Evans RM (май 2014). «Ядерные рецепторы качаются вокруг часов» . Embo сообщает . 15 (5): 518–28. doi : 10.1002/emb.201338271 . PMC 4210094 . PMID 24737872 .
- ^ Jump up to: а беременный Du J, Huang C, Zhou B, Ziegler SF (апрель 2008 г.). «Изоформа-специфическое ингибирование альфа-опосредованной транскрипционной активации ROR FOXP3» . Журнал иммунологии . 180 (7): 4785–92. doi : 10.4049/jimmunol.180.7.4785 . PMID 18354202 .
- ^ Xiong G, Wang C, Evers BM, Zhou BP, Xu R (апрель 2012 г.). «RORα подавляет инвазию опухоли молочной железы, индуцируя экспрессию SEMA3F» . РАНКА . 72 (7): 1728–39. doi : 10.1158/0008-5472.can-11-2762 . PMC 3319846 . PMID 22350413 .
- ^ Paravicini G, Steinmayr M, André E, Becker-André M (октябрь 1996 г.). «Кандидат -кандидат супрессора метастазирования нуклеотидфосфаткиназа NM23 специфически взаимодействует с членами подсемейства ядерного рецептора -сироты ROR/RZR». Биохимическая и биофизическая исследовательская коммуникация . 227 (1): 82–7. doi : 10.1006/bbrc.1996.1471 . PMID 8858107 .
Дальнейшее чтение
[ редактировать ]- Giguère V, Beatty B, Squire J, Copeland NG, Jenkins NA (август 1995). «Ядерный рецептор-сирот ROR Alpha (RORA) отображает в консервативную область гомологии на хромосоме человека 15Q21-Q22 и хромосомы мыши 9». Геномика . 28 (3): 596–8. doi : 10.1006/geno.1995.1197 . PMID 7490103 .
- Steinhilber D, Brungs M, Werz O, Wiesenberg I, Danielsson C, Kahlen JP, et al. (Март 1995 г.). «Ядерный рецептор для мелатонина репрессирует экспрессию гена 5-липоксигеназы в лимфоцитах человека» . Журнал биологической химии . 270 (13): 7037–40. doi : 10.1074/jbc.270.13.7037 . PMID 7706239 .
- Forman BM, Chen J, Blumberg B, Kliewer SA, Henshaw R, Ong ES, et al. (Сентябрь 1994). «Перекрестное разговор между ROR Alpha 1 и семейством Rev-Erb ядерных рецепторов сирот» . Молекулярная эндокринология . 8 (9): 1253–61. doi : 10.1210/mend.8.9.7838158 . PMID 7838158 .
- Беккер-Андре М., Андре Э., Деламартер Дж.Ф. (август 1993). «Идентификация мРНК ядерного рецептора с помощью RT-PCR амплификации консервативных последовательностей мотива цинкового пальца». Биохимическая и биофизическая исследовательская коммуникация . 194 (3): 1371–9. doi : 10.1006/bbrc.1993.1976 . PMID 7916608 .
- Карлберг С., Хофт Ван Хуйсдуйнен Р., Стейпл Дж.К., Деламартер Дж.Ф., Беккер-Андре М. (июнь 1994 г.). «RZRS, новое семейство рецепторов-сирот, связанных с ретиноидом, которые функционируют как мономеры и гомодимеры» . Молекулярная эндокринология . 8 (6): 757–70. doi : 10.1210/mend.8.6.7935491 . PMID 7935491 . S2CID 22342101 .
- Paravicini G, Steinmayr M, André E, Becker-André M (октябрь 1996 г.). «Кандидат -кандидат супрессора метастазирования нуклеотидфосфаткиназа NM23 специфически взаимодействует с членами подсемейства ядерного рецептора -сироты ROR/RZR». Биохимическая и биофизическая исследовательская коммуникация . 227 (1): 82–7. doi : 10.1006/bbrc.1996.1471 . PMID 8858107 .
- Лау П., Бейли П., Доухан Д.Х., Маскат Г.Е. (январь 1999 г.). «Экзогенная экспрессия доминантного отрицательного вектора Roralpha1 в мышечных клетках нарушает дифференцировку: Roralpha1 напрямую взаимодействует с p300 и myod» . Исследование нуклеиновых кислот . 27 (2): 411–20. doi : 10.1093/nar/27.2.411 . PMC 148194 . PMID 9862959 .
- Atkins GB, Hu X, Guenther MG, Rachez C, Freedman LP, Lazar MA (сентябрь 1999 г.). «Коактиваторы для ядерного рецептора сироты Roralpha» . Молекулярная эндокринология . 13 (9): 1550–7. doi : 10.1210/mend.13.9.0343 . PMID 10478845 .
- Meyer T, Kneissel M, Mariani J, Fournier B (август 2000). «Доказательства in vitro и in vivo для ядерного рецептора -сироты Roralpha функции в метаболизме кости» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (16): 9197–202. Bibcode : 2000pnas ... 97.9197M . doi : 10.1073/pnas.150246097 . PMC 16845 . PMID 10900268 .
- Gawlas K, Stunnenberg HG (декабрь 2000 г.). «Дифференциальное связывание и транскрипционное поведение двух высоко связанных рецепторов сирот, ROR Alpha (4) и ROR Beta (1)». Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1494 (3): 236–41. doi : 10.1016/s0167-4781 (00) 00237-2 . PMID 11121580 .
- Delerive P, Chin WW, Suen CS (сентябрь 2002 г.). «Идентификация реверберации (альфа) как нового гена -мишени ROR (альфа)» . Журнал биологической химии . 277 (38): 35013–8. doi : 10.1074/jbc.m202979200 . PMID 12114512 .
- Моретти Р.М., Монтаньяни Марелли М., Мотта М., Лимонта П. (2003). «Роль ядерного рецептора сироты ROR AlphA в контроле метастатического поведения андроген-независимых клеток рака простаты». Онкологические отчеты . 9 (5): 1139–43. doi : 10.3892/или.9.5.1139 . PMID 12168086 .
- Raspè E, Mautino G, Duval C, Fontaine C, Dudez H, Barbier O, et al. (Декабрь 2002 г.). «Транскрипционная регуляция экспрессии гена REV-Erbalpha человека с помощью ядерной рецептор-рецептор-рецептор-рецептор-рецептора, связанной с рецептором сироты, альфа-рецептором сирот» . Журнал биологической химии . 277 (51): 49275–81. doi : 10.1074/jbc.m206215200 . PMID 12377782 .
- Kallen J, Schlaeppi JM, Bitsch F, Delhon I, Fournier B (апрель 2004 г.). «Кристаллическая структура домена связывания лиганда Roralpha Roralpha в комплексе с сульфатом холестерина при 2,2 А» . Журнал биологической химии . 279 (14): 14033–8. doi : 10.1074/jbc.m400302200 . PMID 14722075 .
- Мигита Х, Сатозава Н., Лин Дж. Х., Мордер Дж., Кавай К (январь 2004 г.). «Roralpha1 и Roralpha4 подавляют индуцированную TNF-альфа экспрессию VCAM-1 и ICAM-1 в эндотелиальных клетках человека» . Письма Febs . 557 (1–3): 269–74. Bibcode : 2004febsl.557..269m . doi : 10.1016/s0014-5793 (03) 01502-3 . PMID 14741380 . S2CID 24388280 .
- Мики Н., Икута М., Мацуи Т (апрель 2004 г.). «Индуцированная гипоксией активация гена Alpha4, связанного с рецептором ретиноевой кислоты, путем взаимодействия между индуцируемым гипоксией фактором-1 и SP1» . Журнал биологической химии . 279 (15): 15025–31. doi : 10.1074/jbc.m313186200 . PMID 14742449 .
- Мигита Х., Мордер Дж., Каваи К (март 2004 г.). «Rev-erbalpha активирует NF-Kappab-чувствительные гены в клетках гладких мышц сосудов» . Письма Febs . 561 (1–3): 69–74. Bibcode : 2004febsl.561 ... 69m . doi : 10.1016/s0014-5793 (04) 00118-8 . PMID 15013753 . S2CID 84456190 .
Внешние ссылки
[ редактировать ]- Orphan+ядерный+рецептор+ror-gamma в Национальной библиотеке медицины Медицинской библиотеки США (Mesh)
PDB Галерея |
|---|

