КРЕБ
CREB-TF (CREB, белок, связывающий элемент ответа цАМФ ) [ 1 ] является клеточным фактором транскрипции . Он связывается с определенными ДНК последовательностями называемыми элементами ответа цАМФ (CRE), тем самым увеличивая или уменьшая транскрипцию генов , . [ 2 ] CREB был впервые описан в 1987 году как цАМФ -зависимый фактор транскрипции, регулирующий ген соматостатина . [ 3 ]
Гены, транскрипция которых регулируется CREB, включают: c-fos , BDNF , тирозингидроксилазу , многочисленные нейропептиды (такие как соматостатин , энкефалин , VGF , кортикотропин-рилизинг гормон ), [ 2 ] и гены, участвующие в циркадных часах млекопитающих ( PER1 , PER2 ). [ 4 ]
CREB тесно связан по структуре и функциям с белками CREM ( модулятор элемента ответа цАМФ ) и ATF-1 ( активирующий фактор транскрипции-1 ). Белки CREB экспрессируются у многих животных, включая человека.
CREB играет хорошо документированную роль в пластичности нейронов и формировании долговременной памяти в мозге, и было показано, что он играет важную роль в формировании пространственной памяти . [ 5 ] Снижение уровня регуляции CREB вовлечено в патологию болезни Альцгеймера , и увеличение экспрессии CREB рассматривается как возможная терапевтическая мишень при болезни Альцгеймера. [ 6 ] CREB также играет роль в фотоувлечении у млекопитающих.
Подтипы
[ редактировать ]Следующие гены кодируют CREB или CREB-подобные белки:
- CREB1 ( CREB1 )
- CREB2 переименован в ATF4 ( ATF4 )
- CREB3 ( CREB3 )
- CREB5 ( CREB5 )
- CREB3L1 ( CREB3L1 )
- CREB3L2 ( CREB3L2 )
- CREB3L3 ( CREB3L3 )
- CREB3L4 ( CREB3L4 )
Структура
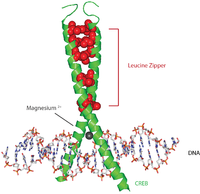
[ редактировать ]Белки CREB активируются путем фосфорилирования различных киназ, включая PKA и Ca. 2+ /кальмодулин-зависимые протеинкиназы по остатку серина 133. [ 7 ] При активации белок CREB привлекает другие коактиваторы транскрипции для связывания с 5'-областью промотора CRE, расположенной выше. Гидрофобные аминокислоты лейцин расположены вдоль внутреннего края альфа-спирали. Эти остатки лейцина прочно связываются с остатками лейцина другого белка CREB, образуя димер. Эта цепочка остатков лейцина образует мотив лейциновой молнии . Белок также имеет ион магния, который облегчает связывание с ДНК.
элемент ответа цАМФ
[ редактировать ]Элемент ответа цАМФ (CRE) является элементом ответа для CREB, который содержит высококонсервативную нуклеотидную последовательность 5'-TGACGTCA-3'. Сайты CRE обычно обнаруживаются выше генов, в областях промотора или энхансера . [ 8 ] В геноме человека насчитывается около 750 000 палиндромных и полусайтовых CRE. Однако большинство этих сайтов остаются несвязанными из-за метилирования цитозина , которое физически препятствует связыванию белка. [ 9 ]
Механизм действия
[ редактировать ]Обобщенная последовательность событий резюмируется следующим образом: сигнал поступает на поверхность клетки, активирует соответствующий рецептор, что приводит к выработке вторичного мессенджера, такого как цАМФ или Са. 2+ , который, в свою очередь, активирует протеинкиназу . Эта протеинкиназа перемещается в ядро клетки , где активирует белок CREB. Активированный белок CREB затем связывается с областью CRE, а затем связывается с CBP (CREB-связывающий белок), который коактивирует его, позволяя ему включать или выключать определенные гены. Связывание ДНК CREB опосредовано через его основной домен лейциновой молнии ( домен bZIP ), как показано на изображении. Данные свидетельствуют о том, что β-адренорецептор ( рецептор, связанный с G-белком ) стимулирует передачу сигналов CREB. [ 10 ]
Функция в мозге
[ редактировать ]CREB выполняет множество функций во многих различных органах, и некоторые из его функций изучались применительно к мозгу. [ 11 ] Считается, что белки CREB в нейронах участвуют в формировании долговременной памяти; [ 12 ] это было показано на морской улитке Aplysia , плодовой мухе Drosophila melanogaster , крысах и мышах (см. CREB в книге «Молекулярное и клеточное познание »). [ 1 ] CREB необходим на поздней стадии долговременной потенциации . CREB также играет важную роль в развитии наркозависимости и тем более психологической зависимости . [ 13 ] [ 14 ] [ 15 ] Существуют активаторная и репрессорная формы CREB. Мухи, генетически модифицированные для сверхэкспрессии неактивной формы CREB, теряют способность сохранять долговременную память. CREB также важен для выживания нейронов, как было показано на генно-инженерных мышах, у которых CREB и CREM были удалены в мозге. Если CREB теряется во всем развивающемся эмбрионе мыши, мыши умирают сразу после рождения, что еще раз подчеркивает критическую роль CREB в обеспечении выживания нейронов.
Связь с болезнями
[ редактировать ]Нарушение функции CREB в головном мозге может способствовать развитию и прогрессированию болезни Хантингтона .
Аномалии белка, который взаимодействует с доменом KID CREB, CREB-связывающего белка (CBP), связаны с синдромом Рубинштейна-Тайби .
Есть некоторые данные, позволяющие предположить, что недостаточное функционирование CREB связано с большим депрессивным расстройством . [ 16 ] Крысы с депрессией со сверхэкспрессией CREB в зубчатой извилине вели себя аналогично крысам, получавшим антидепрессанты. [ 17 ] Посмертные исследования также показали, что кора головного мозга пациентов с нелеченым большим депрессивным расстройством содержит пониженные концентрации CREB по сравнению как со здоровыми людьми из контрольной группы, так и с пациентами, получавшими антидепрессанты. [ 17 ] Функция CREB может модулироваться с помощью сигнального пути, возникающего в результате связывания серотонина и норадреналина с постсинаптическими рецепторами, связанными с G-белком. Дисфункция этих нейротрансмиттеров также связана с большим депрессивным расстройством. [ 16 ]
Также считается, что CREB участвует в росте некоторых видов рака.
Участие в циркадных ритмах
[ редактировать ]Смещение циркадных часов млекопитающих устанавливается посредством световой индукции PER . Свет возбуждает меланопсин содержащие фоточувствительные ганглиозные клетки сетчатки , которые передают сигнал супрахиазматическому ядру (SCN) через ретиногипоталамический тракт (RHT). Возбуждение RHT сигнализирует о высвобождении глутамата, который воспринимается NMDA-рецепторами на SCN, что приводит к притоку кальция в SCN. Кальций индуцирует активность Ca 2+ / кальмодулин-зависимые протеинкиназы , что приводит к активации PKA , PKC и CK2 . [ 18 ] Эти киназы затем фосфорилируют CREB циркадным образом, что дополнительно регулирует экспрессию генов. [ 19 ] Фосфорилированный CREB распознает элемент ответа цАМФ и служит фактором транскрипции для Per1 и Per2 , двух генов, которые регулируют циркадные часы млекопитающих. Эта индукция белка PER может вовлечь циркадные часы в циклы света/темноты, ингибируя собственную транскрипцию через петлю обратной связи транскрипции-трансляции, которая может опережать или задерживать циркадные часы. Однако реакция на индукцию белков PER1 и PER2 значима только в течение субъективной ночи. [ 4 ]
Открытие участия CREB в циркадных ритмах
[ редактировать ]Майкл Гринберг впервые продемонстрировал роль CREB в циркадных часах млекопитающих в 1993 году посредством серии экспериментов, которые коррелировали фазоспецифические световые импульсы с фосфорилированием CREB. In vitro свет в ночное время увеличивал фосфорилирование CREB, а не уровни белка CREB. In vivo световые импульсы, индуцирующие фазовый сдвиг, во время субъективной ночи коррелируют с фосфорилированием CREB в SCN. [ 20 ] Эксперименты Гюнтера Шютца в 2002 году продемонстрировали, что мутантные мыши, у которых отсутствует сайт фосфорилирования Ser142, не способны индуцировать ген регулятора часов mPer1 в ответ на световой импульс. Более того, эти мутантные мыши с трудом приспосабливались к циклам света и темноты. [ 21 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Бурчуладзе; и др. (1994). «Дефицит долговременной памяти у мышей с целевой мутацией белка, связывающего элемент, реагирующий на цАМФ». Клетка . 79 (1): 59–68. дои : 10.1016/0092-8674(94)90400-6 . ПМИД 7923378 . S2CID 17250247 .
- ^ Jump up to: а б Первс, Дейл; Джордж Дж. Августин; Дэвид Фицпатрик; Уильям К. Холл; Энтони-Сэмюэл ЛаМантиа; Джеймс О. Макнамара и Леонард Э. Уайт (2008). Нейронаука (4-е изд.). Синауэр Ассошиэйтс. стр. 170–6. ISBN 978-0-87893-697-7 .
- ^ Монтмини, MR; Билезикджян, Л.М. (1987). «Связывание ядерного белка с элементом ответа циклического АМФ гена соматостатина». Природа . 328 (6126): 175–178. Бибкод : 1987Natur.328..175M . дои : 10.1038/328175a0 . ПМИД 2885756 . S2CID 4345292 .
- ^ Jump up to: а б Дибнер, Чарна; Шиблер, Ули; Альбрехт, Урс (2010). «Циркадная система синхронизации млекопитающих: организация и координация центральных и периферических часов» (PDF) . Ежегодный обзор физиологии . 72 (1): 517–549. doi : 10.1146/annurev-psyol-021909-135821 . ПМИД 20148687 .
- ^ Сильва; и др. (1998). «КРЭБ и память» (PDF) . Ежегодный обзор неврологии . 21 : 127–148. дои : 10.1146/annurev.neuro.21.1.127 . ПМИД 9530494 . Архивировано из оригинала (PDF) 28 августа 2008 года . Проверено 22 января 2010 г.
- ^ Снижение экспрессии CREB в мозге, страдающем болезнью Альцгеймера, и в нейронах гиппокампа крыс, обработанных АБ.
- ^ Шайвиц, Адам Дж.; Гринберг, Майкл Э. (1999). «CREB: индуцированный стимулом фактор транскрипции, активируемый разнообразным набором внеклеточных сигналов». Ежегодный обзор биохимии . 68 (1): 821–861. doi : 10.1146/annurev.biochem.68.1.821 . ПМИД 10872467 .
- ^ Карлесон, Вашингтон; Думан, РС; Нестлер, Э.Дж. (август 2005 г.). «Многоликий CREB». Тенденции в нейронауках . 28 (8): 436–445. doi : 10.1016/j.tins.2005.06.005 . ПМИД 15982754 . S2CID 6480593 .
- ^ Альтарехос, Джудит Ю.; Монтмини, Марк (март 2011 г.). «CREB и коактиваторы CRTC: сенсоры гормональных и метаболических сигналов» . Nature Reviews Молекулярно-клеточная биология . 12 (3): 141–151. дои : 10.1038/nrm3072 . ISSN 1471-0072 . ПМЦ 4324555 . ПМИД 21346730 .
- ^ Пирс, Александр; Сандерс, Люси; Брайтон, Пол Дж.; Рана, Шаши; Конье, Джастин С.; Уиллетс, Джонатон М. (1 октября 2017 г.). «Взаимная регуляция передачи сигналов белка, связывающего ответный элемент цАМФ, активируемого β2-адренорецептором, с помощью аррестина2 и аррестина3» (PDF) . Сотовая сигнализация . 38 : 182–191. дои : 10.1016/j.cellsig.2017.07.011 . ISSN 0898-6568 . ПМИД 28733084 .
- ^ Карлесон В.А., Думан Р.С., Нестлер Э.Дж. (август 2005 г.). «Многоликий CREB». Тенденции в нейронауках . 28 (8): 436–45. doi : 10.1016/j.tins.2005.06.005 . ПМИД 15982754 . S2CID 6480593 .
- ^ Кандел, Эрик Р. (14 мая 2012 г.). «Молекулярная биология памяти: цАМФ, PKA, CRE, CREB-1, CREB-2 и CPEB» . Молекулярный мозг . 5:14 . дои : 10.1186/1756-6606-5-14 . ISSN 1756-6606 . ПМК 3514210 . ПМИД 22583753 .
- ^ Назарян А., Сунь В.Л., Чжоу Л., Кемен Л.М., Дженаб С., Хиноны-Дженаб В. (апрель 2009 г.). «Половые различия в базальных и кокаин-индуцированных изменениях белков PKA и CREB в прилежащем ядре». Психофармакология . 203 (3): 641–50. дои : 10.1007/s00213-008-1411-5 . ПМИД 19052730 . S2CID 24064950 .
- ^ Ван Ю, Гецци А., Инь Дж. К., Аткинсон Н. С. (июнь 2009 г.). «Регуляция CREB экспрессии генов BK-каналов лежит в основе быстрой толерантности к лекарствам» . Гены, мозг и поведение . 8 (4): 369–76. дои : 10.1111/j.1601-183X.2009.00479.x . ПМЦ 2796570 . ПМИД 19243452 .
- ^ ДиРокко Д.П., Шайнер З.С., Синдреу К.Б., Чан Г.К., Шторм Д.Р. (февраль 2009 г.). «Роль стимулированных кальмодулином аденилатциклаз в сенсибилизации кокаина» . Журнал неврологии . 29 (8): 2393–403. doi : 10.1523/JNEUROSCI.4356-08.2009 . ПМК 2678191 . ПМИД 19244515 .
- ^ Jump up to: а б Белмейкер, Р.Х.; Агам, Галила (2008). «Большое депрессивное расстройство». Медицинский журнал Новой Англии . 358 (1): 55–68. дои : 10.1056/nejmra073096 . ПМИД 18172175 .
- ^ Jump up to: а б Бленди, JA (2006). «Роль CREB в депрессии и лечении антидепрессантами». Биологическая психиатрия . 59 (12): 1144–50. doi : 10.1016/j.biopsych.2005.11.003 . ПМИД 16457782 . S2CID 20918484 .
- ^ Айер, Раджашекар; Ван, Тунфэй; Джилетт, Марта (19 сентября 2014 г.). «Циркадные ворота функциональности нейронов: основа итеративной метапластичности» . Границы системной нейронауки . 8 : 164. дои : 10.3389/fnsys.2014.00164 . ПМК 4168688 . ПМИД 25285070 .
- ^ Обриетан, Карл; Импи, Сорен; Смит, Дэйв; Атос, Хайме; Шторм, Деррик Р. (11 апреля 2002 г.). «Циркадная регуляция экспрессии генов, опосредованной элементом ответа цАМФ, в супрахиазматических ядрах» . Журнал биологической химии . 274 (25): 17748–17756. дои : 10.1074/jbc.274.25.17748 . ПМИД 10364217 .
- ^ Джинти, Д.Д.; Корнхаузер, Дж. М.; Томпсон, Массачусетс; Бадинг, Х.; Мэйо, Кентукки; Такахаши, Дж.С.; Гринберг, Мэн (9 апреля 1993 г.). «Регуляция фосфорилирования CREB в супрахиазматическом ядре с помощью света и циркадных часов». Наука . 260 (5105): 238–241. Бибкод : 1993Sci...260..238G . дои : 10.1126/science.8097062 . ISSN 0036-8075 . ПМИД 8097062 .
- ^ Гау, Дэниел; Лембергер, Томас; фон Галль, Шарлотта; Крец, Оливер; Ле Минь, Нгует; Гасс, Питер; Шмид, Вольфганг; Шиблер, Ули; Корф, Хорст В. (11 апреля 2002 г.). «Фосфорилирование CREB Ser142 регулирует индуцированные светом фазовые сдвиги циркадных часов» . Нейрон . 34 (2): 245–253. дои : 10.1016/S0896-6273(02)00656-6 . ПМИД 11970866 . S2CID 14507897 .
- Библиография
- Лорен Слейтер (2005). Открывая ящик Скиннера: великие психологические эксперименты двадцатого века . Нью-Йорк: WW Norton & Company. ISBN 978-0-393-32655-0 .
- Барко А., Бейли С., Кандел Э. (2006). «Общие молекулярные механизмы в явной и неявной памяти» . Дж. Нейрохем . 97 (6): 1520–33. дои : 10.1111/j.1471-4159.2006.03870.x . ПМИД 16805766 .
- Конкрайт М., Монтмини М. (2005). «CREB: сообщник рака, которому не предъявлено обвинение». Тенденции клеточной биологии . 15 (9): 457–9. дои : 10.1016/j.tcb.2005.07.007 . ПМИД 16084096 .
- Мантамадиотис Т., Лембергер Т., Блекманн С., Керн Х., Крец О., Мартин Вильяльба А., Тронш Ф., Келлендонк С., Гау Д., Капфхаммер Дж., Отто С., Шмид В., Шютц Г. (2002). «Нарушение функции CREB в головном мозге приводит к нейродегенерации» . Нат. Жене . 31 (1): 47–54. дои : 10.1038/ng882 . ПМИД 11967539 . S2CID 22014116 .
- Майр Б., Монтмини М. (2001). «Регуляция транскрипции с помощью фосфорилирования-зависимого фактора CREB». Нат. Преподобный мол. Клеточная Биол . 2 (8): 599–609. дои : 10.1038/35085068 . ПМИД 11483993 . S2CID 1056720 .
- Инь Дж., Дель Веккио М., Чжоу Х., Талли Т. (1995). «CREB как модулятор памяти: индуцированная экспрессия изоформы активатора dCREB2 улучшает долговременную память у дрозофилы» . Клетка . 81 (1): 107–15. дои : 10.1016/0092-8674(95)90375-5 . ПМИД 7720066 . S2CID 15863948 .
- Инь Дж., Уоллак Дж., Дель Веккио М., Уайлдер Э., Чжоу Х., Куинн В., Талли Т. (1994). «Индукция доминантно-негативного трансгена CREB специфически блокирует долговременную память у дрозофилы». Клетка . 79 (1): 49–58. дои : 10.1016/0092-8674(94)90399-9 . ПМИД 7923376 . S2CID 33623585 .
Внешние ссылки
[ редактировать ]- http://www.ebi.ac.uk/interpro/entry/IPR001630
- Йоханнессен М., Педерсен Делганди М. и Моенс У. (2004) – Что возбуждает CREB? - Сигнал сотовой связи.; 10:1211-1227. https://web.archive.org/web/20070928090058/http://www.sigtrans.org/publications/what-turns-creb-on/
- https://web.archive.org/web/20060902183214/http://focus.hms.harvard.edu//2001/Oct26_2001/neuroscience.html
- CREB + Белок в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- дрозофилы Белок А, связывающий элемент ответа циклического АМФ - The Interactive Fly
- дрозофилы Белок B, связывающий элемент ответа циклического АМФ в положении 17А - The Interactive Fly