Эстроген
| Эстроген | |
|---|---|
| Класс препарата | |
 Эстрадиол , основной половой гормон эстрогена у человека и широко используемый препарат. | |
| Идентификаторы классов | |
| Использовать | Контрацепция , менопауза , гипогонадизм , трансгендерные женщины , рак простаты , рак молочной железы и др. |
| код АТС | G03C |
| Биологическая цель | Рецепторы эстрогена ( ERα , ERβ , mER (например, GPER и др.)) |
| Внешние ссылки | |
| МеШ | D004967 |
| Юридический статус | |
| В Викиданных | |
Эстроген ( британский английский : эстроген ; см. различия в написании ) — категория половых гормонов, ответственных за развитие и регуляцию женской репродуктивной системы и вторичных половых признаков . [1] [2] Существует три основных эндогенных эстрогена, обладающих эстрогенной гормональной активностью: эстрон (Е1), эстрадиол (Е2) и эстриол (Е3). [1] [3] Эстрадиол, эстран , является наиболее мощным и распространенным. [1] Другой эстроген, называемый эстролом (Е4), вырабатывается только во время беременности.
Эстрогены синтезируются у всех позвоночных. [4] и некоторые насекомые. [5] Количественно эстрогены циркулируют на более низких уровнях, чем андрогены, как у мужчин, так и у женщин. [6] Хотя уровень эстрогена у мужчин значительно ниже, чем у женщин, тем не менее, эстрогены играют важную физиологическую роль у мужчин. [7]
Как и все стероидные гормоны , эстрогены легко диффундируют через клеточную мембрану . Попав внутрь клетки, они связываются и активируют рецепторы эстрогена (ER), которые, в свою очередь модулируют экспрессию , многих генов . [8] Кроме того, эстрогены связываются с мембранными рецепторами эстрогена (mER) и активируют их. [9] [10] например GPER (GPR30). [11]
Помимо своей роли естественных гормонов, эстрогены используются в качестве лекарств , например, в менопаузальной гормональной терапии , гормональном контроле над рождаемостью и феминизирующей гормональной терапии для трансгендерных женщин , интерсекс-людей и небинарных людей .
Синтетические и природные эстрогены были обнаружены в окружающей среде и называются ксеноэстрогенами . Эстрогены относятся к широкому спектру соединений, нарушающих работу эндокринной системы (EDC), и могут вызывать проблемы со здоровьем и репродуктивную дисфункцию как у диких животных, так и у людей. [12] [13]
Типы и примеры
[ редактировать ] Структуры основных эндогенных эстрогенов |
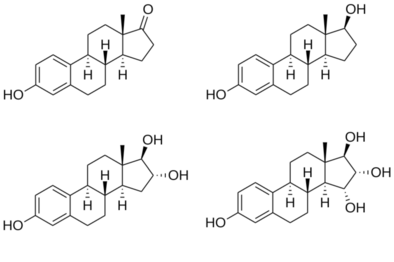
Четырьмя основными природными эстрогенами являются эстрон (Е1), эстрадиол (Е2), эстриол (Е3) и эстетрол (Е4). Эстрадиол (Е2) является преобладающим эстрогеном в репродуктивном возрасте как с точки зрения абсолютного уровня в сыворотке, так и с точки зрения эстрогенной активности. Во время менопаузы эстрон является преобладающим циркулирующим эстрогеном, а во время беременности эстриол является преобладающим циркулирующим эстрогеном с точки зрения уровня сыворотки. При подкожной инъекции мышам эстрадиол примерно в 10 раз более эффективен, чем эстрон, и примерно в 100 раз более эффективен, чем эстриол. [14] Таким образом, эстрадиол является наиболее важным эстрогеном у небеременных женщин, находящихся между менархе стадиями жизни и менопаузы. Однако во время беременности эта роль переходит к эстриолу, а в постменопаузе эстрон становится основной формой эстрогена в организме. Другой тип эстрогена, называемый эстролом (Е4), вырабатывается только во время беременности. Все различные формы эстрогена синтезируются из андрогенов , особенно тестостерона и андростендиона , с помощью фермента ароматазы .
К минорным эндогенным эстрогенам, в биосинтезе которых не участвует ароматаза , относятся 27-гидроксихолестерин , дегидроэпиандростерон (ДГЭА), 7-оксо-ДГЭА , 7α-гидрокси-ДГЭА , 16α-гидрокси-ДГЭА , 7β-гидроксиэпиандростерон , андростендион (А4), андростенедиол (А5), 3α-андростандиол и 3β-андростандиол . [15] [16] Некоторые метаболиты эстрогена, такие как катехоловые эстрогены 2-гидроксиэстрадиол , 2-гидроксиэстрон , 4-гидроксиэстрадиол и 4-гидроксиэстрон , а также 16α-гидроксиэстрон , также являются эстрогенами с различной степенью активности. [17] Биологическое значение этих второстепенных эстрогенов не совсем ясно.
Биологическая функция
[ редактировать ]
Действие эстрогена опосредовано рецептором эстрогена (ER), димерным ядерным белком, который связывается с ДНК и контролирует экспрессию генов . Как и другие стероидные гормоны, эстроген пассивно проникает в клетку, где связывается и активирует рецептор эстрогена. Комплекс эстроген:ER связывается со специфическими последовательностями ДНК, называемыми элементом гормонального ответа , для активации транскрипции генов-мишеней (в исследовании с использованием эстроген-зависимой линии клеток рака молочной железы в качестве модели было идентифицировано 89 таких генов). [19] Поскольку эстроген проникает во все клетки, его действие зависит от присутствия ЭР в клетке. ЭР экспрессируется в определенных тканях, включая яичник, матку и молочную железу. Метаболические эффекты эстрогена у женщин в постменопаузе связаны с генетическим полиморфизмом ЭР. [20]
Хотя эстрогены присутствуют как у мужчин , так и у женщин , у женщин репродуктивного возраста они обычно присутствуют в значительно более высоких уровнях. Они способствуют развитию женских вторичных половых признаков , таких как увеличение груди , потемнение и увеличение сосков , [21] утолщение эндометрия и другие аспекты регуляции менструального цикла. У мужчин эстроген регулирует определенные функции репродуктивной системы, важные для созревания сперматозоидов. [22] [23] [24] и может быть необходим для здорового либидо . [25]
| Эстроген | Подсказка ER Подсказка RBA (%) | Вес матки (%) | Утеротрофия | Подсказка по ЛГ (%) | SHBG RBA (%) |
|---|---|---|---|---|---|
| Контроль | – | 100 | – | 100 | – |
| Эстрадиол (E2) | 100 | 506 ± 20 | +++ | 12–19 | 100 |
| Эстрон (E1) | 11 ± 8 | 490 ± 22 | +++ | ? | 20 |
| Эстриол (Е3) | 10 ± 4 | 468 ± 30 | +++ | 8–18 | 3 |
| Эстетрол (Е4) | 0.5 ± 0.2 | ? | Неактивный | ? | 1 |
| 17α-эстрадиол | 4.2 ± 0.8 | ? | ? | ? | ? |
| 2-гидроксиэстрадиол | 24 ± 7 | 285 ± 8 | + б | 31–61 | 28 |
| 2-метоксиэстрадиол | 0.05 ± 0.04 | 101 | Неактивный | ? | 130 |
| 4-гидроксиэстрадиол | 45 ± 12 | ? | ? | ? | ? |
| 4-метоксиэстрадиол | 1.3 ± 0.2 | 260 | ++ | ? | 9 |
| 4-фторэстрадиол а | 180 ± 43 | ? | +++ | ? | ? |
| 2-гидроксиэстрон | 1.9 ± 0.8 | 130 ± 9 | Неактивный | 110–142 | 8 |
| 2-метоксиэстрон | 0.01 ± 0.00 | 103 ± 7 | Неактивный | 95–100 | 120 |
| 4-гидроксиэстрон | 11 ± 4 | 351 | ++ | 21–50 | 35 |
| 4-метоксиэстрон | 0.13 ± 0.04 | 338 | ++ | 65–92 | 12 |
| 16α-гидроксиэстрон | 2.8 ± 1.0 | 552 ± 42 | +++ | 7–24 | <0,5 |
| 2-гидроксистриол | 0.9 ± 0.3 | 302 | + б | ? | ? |
| 2-метоксиэстриол | 0.01 ± 0.00 | ? | Неактивный | ? | 4 |
| Примечания: Значения являются средними ± SD или диапазоном. ER RBA = Относительное сродство связывания с эстрогеновыми рецепторами крысы матки цитозоля . Масса матки = процентное изменение сырой массы матки у крыс с удаленными яичниками через 72 часа при непрерывном введении 1 мкг/час с помощью подкожно имплантированных осмотических насосов . Уровни ЛГ = уровни лютеинизирующего гормона относительно исходного уровня у крыс с удаленными яичниками после 24–72 часов непрерывного введения через подкожный имплантат. Сноски: а = Синтетический (т.е. не эндогенный ). б = Атипичный утеротрофический эффект, который достигает плато в течение 48 часов (утеротрофия эстрадиола продолжается линейно до 72 часов). Источники: см. шаблон. | |||||
Обзор действий
[ редактировать ]- Опорно-двигательный аппарат
- Анаболическое действие : увеличивает мышечную массу и силу, скорость регенерации мышц и плотность костей , повышает чувствительность к физическим нагрузкам, защищает от повреждения мышц, усиливает синтез коллагена , увеличивает содержание коллагена в соединительных тканях , сухожилиях и связках , а также снижает жесткость сухожилий. и связки (особенно во время менструации ). Снижение жесткости сухожилий снижает предрасположенность женщин к мышечным напряжениям, но мягкие связки гораздо более подвержены травмам ( разрывы передней крестообразной связки в 2–8 раз чаще встречаются у женщин, чем у мужчин). [26] [27] [28] [29]
- Уменьшает резорбцию кости , увеличивает костеобразование
- Было показано, что у мышей эстроген увеличивает долю самых быстросокращающихся мышечных волокон (тип IIX) более чем на 40%. [30]
- Метаболический
- Противовоспалительные свойства
- Ускорить метаболизм
- Распределение гиноидного жира : увеличение накопления жира или эстрогенного жира в некоторых частях тела, таких как грудь, ягодицы и ноги, но уменьшение количества брюшного и висцерального жира (андрогенное ожирение). [31] [32] [33]
- Эстрадиол также регулирует расход энергии, гомеостаз массы тела и, по-видимому, оказывает гораздо более сильное воздействие на ожирение, чем тестостерон в целом. [34]
- Другие структурные
- Уход за сосудами и кожей
- белка Синтез
- Увеличение выработки печенью . связывающих белков
- Увеличивает выработку гепатокина адропина . [35]
- Коагуляция
- Повышение уровня циркулирующих факторов 2 , 7 , 9 , 10 , плазминогена.
- Снижение антитромбина III
- Увеличение тромбоцитов адгезии
- Увеличение фактора Виллебранда (эстроген -> ангиотензин II -> вазопрессин )
- Увеличение PAI-1 и PAI-2 также за счет ангиотензина II.
- Липид
- Увеличение ЛПВП , триглицеридов
- Снижение ЛПНП , отложение жира
- Баланс жидкости
- Задержка соли ( натрия ) и воды, включая отек и отек лица. [36] [37]
- Эстроген связан с отеками , включая отек лица и живота.
- Меланин
- Известно, что эстроген вызывает потемнение кожи, особенно на лице и ареолах . [38] У женщин с бледной кожей во время беременности кожа становится более коричневой и желтой в результате увеличения уровня эстрогена, известного как «маска беременности» . [39] Эстроген может объяснить, почему у женщин глаза темнее, чем у мужчин, а также меньший риск рака кожи, чем у мужчин; Европейское исследование показало, что у женщин обычно более темная кожа, чем у мужчин. [40] [41]
- Функция легких
- Сексуальный
- Опосредованное формирование женских вторичных половых признаков
- Стимулировать эндометрия рост
- Увеличение матки роста
- Увеличение вагинальной смазки
- Утолщает влагалища стенку
- матки Слизистая оболочка
- Эстроген вместе с прогестероном способствует и поддерживает слизистую оболочку матки при подготовке к имплантации оплодотворенной яйцеклетки и поддержании функции матки в период беременности, а также активирует рецептор окситоцина в миометрии.
- Овуляция
- Скачок уровня эстрогена вызывает выброс лютеинизирующего гормона , который затем вызывает овуляцию, высвобождая яйцеклетку из граафова фолликула в яичнике .
- Сексуальное поведение
- Эстроген необходим самкам млекопитающих для проявления лордоза во время течки (когда у животных «течка»). [43] [44] и регулируется вентромедиальным ядром гипоталамуса Такое поведение необходимо для сексуальной восприимчивости у этих млекопитающих . [45]
- Половое влечение зависит от андрогенов уровня [46] только в присутствии эстрогена, но без эстрогена уровень свободного тестостерона фактически снижает сексуальное желание (вместо увеличения полового влечения), как показано на примере тех женщин, у которых гипоактивное расстройство полового влечения , и сексуальное желание у этих женщин можно восстановить путем введения. эстрогена (при использовании оральных контрацептивов). [47]
Женское половое развитие
[ редактировать ]Эстрогены ответственны за развитие женских вторичных половых признаков в период полового созревания , включая развитие груди , расширение бедер и распределение жира у женщин . И наоборот, андрогены ответственны за на лобке и волос рост теле , а также за прыщи и неприятный запах подмышками .
Развитие груди
[ редактировать ]Эстроген в сочетании с гормоном роста (ГР) и его секреторным продуктом инсулиноподобным фактором роста 1 (ИФР-1) играет решающую роль в обеспечении развития груди в период полового созревания , а также в созревании груди во время беременности при подготовке к лактации и грудному вскармливанию . [48] [49] Эстроген несет главную и прямую ответственность за индуцирование протокового компонента развития молочной железы. [50] [51] [52] а также вызывает отложение жира и рост соединительной ткани . [50] [51] Он также косвенно участвует в лобулоальвеолярном компоненте за счет увеличения экспрессии рецепторов прогестерона в молочной железе. [50] [52] [53] и индуцируя секрецию пролактина . [54] [55] и пролактин , допускаемые эстрогеном, Прогестерон действуют вместе, обеспечивая завершение лобулоальвеолярного развития во время беременности. [51] [56]
Андрогены , такие как тестостерон, мощно противодействуют действию эстрогена в молочных железах, например, путем снижения экспрессии в них рецепторов эстрогена . [57] [58]
Женская репродуктивная система
[ редактировать ]Эстрогены отвечают за созревание и поддержание влагалища и матки , а также участвуют в функции яичников , например, в созревании фолликулов яичников . Кроме того, эстрогены играют важную роль в регуляции гонадотропинов секреции . По этим причинам эстрогены необходимы для женской фертильности .
Нейропротекция и восстановление ДНК
[ редактировать ], регулируемые эстрогенами, репарации ДНК Механизмы в головном мозге оказывают нейропротекторное действие. [59] Эстроген регулирует транскрипцию ДНК генов эксцизионной репарации оснований , а также транслокацию ферментов эксцизионной репарации оснований между различными субклеточными компартментами.
Мозг и поведение
[ редактировать ]Секс-драйв
[ редактировать ]Эстрогены участвуют в либидо (половом влечении) как у женщин, так и у мужчин.
Познание
[ редактировать ]Оценка вербальной памяти часто используется как один из показателей более высокого уровня познания . Эти показатели варьируются прямо пропорционально уровням эстрогена на протяжении менструального цикла, беременности и менопаузы. Кроме того, эстрогены, вводимые вскоре после естественной или хирургической менопаузы, предотвращают снижение вербальной памяти. Напротив, эстрогены мало влияют на вербальную память, если их впервые вводить спустя годы после менопаузы. [60] Эстрогены также оказывают положительное влияние на другие показатели когнитивной функции. [61] Однако влияние эстрогенов на когнитивные функции не всегда благоприятно и зависит от времени приема дозы и типа измеряемых когнитивных навыков. [62]
Защитное воздействие эстрогенов на когнитивные функции может быть опосредовано противовоспалительным действием эстрогена на мозг. [63] Исследования также показали, что ген аллеля Met и уровень эстрогена опосредуют эффективность задач рабочей памяти, зависящих от префронтальной коры . [64] [65] Исследователи призвали к дальнейшим исследованиям, чтобы пролить свет на роль эстрогена и его потенциал для улучшения когнитивных функций. [66]
Психическое здоровье
[ редактировать ]женщин Считается, что эстроген играет значительную роль в психическом здоровье . Внезапная отмена эстрогена, колебания эстрогена и периоды устойчивого низкого уровня эстрогена коррелируют со значительным снижением настроения. клиническое выздоровление от послеродовой , перименопаузальной и постменопаузальной депрессии эффективно после стабилизации и/или восстановления уровня эстрогена. Было показано, что [67] [68] [69] Обострение менструального цикла (включая менструальный психоз) обычно провоцируется низким уровнем эстрогена. [70] и часто ошибочно принимают за предменструальное дисфорическое расстройство . [71]
Компульсии у самцов лабораторных мышей, например, при обсессивно-компульсивном расстройстве (ОКР), могут быть вызваны низким уровнем эстрогена. Когда уровень эстрогена повышался за счет увеличения активности фермента ароматазы у самцов лабораторных мышей, количество ритуалов ОКР резко уменьшалось. Уровни гипоталамического белка в гене COMT повышаются за счет повышения уровня эстрогена, который, как полагают, возвращает мышей, которые демонстрировали ритуалы ОКР, к нормальной активности. В конечном итоге подозревается дефицит ароматазы, которая участвует в синтезе эстрогена у людей и имеет терапевтическое значение у людей, страдающих обсессивно-компульсивным расстройством. [72]
Было показано, что местное применение эстрогена в гиппокампе крыс ингибирует обратный захват серотонина . Напротив, было показано, что местное применение эстрогена блокирует способность флувоксамина замедлять клиренс серотонина, что позволяет предположить, что на те же пути, которые участвуют в эффективности СИОЗС, также могут влиять компоненты местных сигнальных путей эстрогена. [73]
Родительство
[ редактировать ]Исследования также показали, что у отцов был более низкий уровень кортизола и тестостерона, но более высокий уровень эстрогена (эстрадиола), чем у неотцов. [74]
Переедание
[ редактировать ]Эстроген может играть роль в подавлении переедания . Заместительная гормональная терапия с использованием эстрогена может быть возможным лечением переедания у женщин. Было показано, что замена эстрогена подавляет переедание у самок мышей. [75] Механизм, с помощью которого замена эстрогена подавляет переедание, включает замену нейронов серотонина (5-НТ). Обнаружено, что у женщин, демонстрирующих переедание, повышенное поглощение мозгом нейрона 5-HT и, следовательно, меньшее количество нейромедиатора серотонина в спинномозговой жидкости. [76] Эстроген активирует нейроны 5-HT, что приводит к подавлению переедания, например пищевого поведения. [75]
Также предполагается, что существует взаимодействие между уровнем гормонов и питанием в разные моменты женского менструального цикла . Исследования предсказывают повышенное эмоциональное переедание во время гормональных изменений, которые характеризуются высокими уровнями прогестерона и эстрадиола , возникающими в середине лютеиновой фазы . Предполагается, что эти изменения происходят из-за изменений мозга в течение менструального цикла, которые, вероятно, являются геномным эффектом гормонов. Эти эффекты вызывают изменения менструального цикла, которые приводят к выбросу гормонов, что приводит к изменениям в поведении, особенно к перееданию и эмоциональному перееданию. Это особенно заметно среди женщин, которые генетически уязвимы к фенотипам переедания. [77]
Переедание связано со снижением эстрадиола и увеличением прогестерона. [78] Кламп и др. [79] Прогестерон может смягчать эффекты низкого уровня эстрадиола (например, при нарушенном пищевом поведении), но это может быть верно только для женщин, у которых были клинически диагностированы эпизоды переедания (ПБ). Нерегулируемое питание более тесно связано с такими гормонами яичников у женщин с ПБ, чем у женщин без ПБ. [79]
Имплантация гранул 17β-эстрадиола мышам с удаленными яичниками значительно снизила склонность к перееданию, а инъекции GLP-1 мышам с удаленными яичниками снизили склонность к перееданию. [75]
Связь между перееданием, фазой менструального цикла и гормонами яичников коррелирует. [78] [80] [81]
Маскулинизация у грызунов
[ редактировать ]У грызунов эстрогены (которые локально ароматизируются из андрогенов в мозге) играют важную роль в психосексуальной дифференциации, например, путем маскулинизации территориального поведения; [82] то же самое нельзя сказать и о людях. [83] У людей маскулинизирующее воздействие пренатальных андрогенов на поведение (и другие ткани, за возможным исключением воздействия на кости), по-видимому, действует исключительно через андрогенные рецепторы. [84] Следовательно, полезность моделей на грызунах для изучения психосексуальной дифференциации человека была поставлена под сомнение. [85]
Скелетная система
[ редактировать ]Эстрогены ответственны как за скачок пубертатного роста, который вызывает ускорение линейного роста, так и за закрытие эпифизов , которое ограничивает рост и длину конечностей как у женщин, так и у мужчин. Кроме того, эстрогены отвечают за созревание костей и поддержание минеральной плотности костей на протяжении всей жизни. Из-за гипоэстрогении риск остеопороза увеличивается во время менопаузы .
Сердечно-сосудистая система
[ редактировать ]Женщины меньше страдают от сердечно-сосудистых заболеваний из-за сосудозащитного действия эстрогена, который помогает предотвратить атеросклероз. [86] Это также помогает поддерживать тонкий баланс между борьбой с инфекциями и защитой артерий от повреждений, тем самым снижая риск сердечно-сосудистых заболеваний. [87] Во время беременности высокий уровень эстрогенов увеличивает свертываемость крови и риск венозной тромбоэмболии . Было показано, что эстроген усиливает активность пептидного гормона адропина . [35]
| Абсолютная частота возникновения первой ВТЭ на 10 000 человеко-лет во время беременности и в послеродовом периоде | ||||||||
|---|---|---|---|---|---|---|---|---|
| Шведские данные А | Шведские данные B | Английские данные | Датские данные | |||||
| Период времени | Н | Ставка (95% ДИ) | Н | Ставка (95% ДИ) | Н | Ставка (95% ДИ) | Н | Ставка (95% ДИ) |
| Вне беременности | 1105 | 4.2 (4.0–4.4) | 1015 | 3.8 (?) | 1480 | 3.2 (3.0–3.3) | 2895 | 3.6 (3.4–3.7) |
| Дородовый период | 995 | 20.5 (19.2–21.8) | 690 | 14.2 (13.2–15.3) | 156 | 9.9 (8.5–11.6) | 491 | 10.7 (9.7–11.6) |
| Триместр 1 | 207 | 13.6 (11.8–15.5) | 172 | 11.3 (9.7–13.1) | 23 | 4.6 (3.1–7.0) | 61 | 4.1 (3.2–5.2) |
| Триместр 2 | 275 | 17.4 (15.4–19.6) | 178 | 11.2 (9.7–13.0) | 30 | 5.8 (4.1–8.3) | 75 | 5.7 (4.6–7.2) |
| Триместр 3 | 513 | 29.2 (26.8–31.9) | 340 | 19.4 (17.4–21.6) | 103 | 18.2 (15.0–22.1) | 355 | 19.7 (17.7–21.9) |
| Вокруг доставки | 115 | 154.6 (128.8–185.6) | 79 | 106.1 (85.1–132.3) | 34 | 142.8 (102.0–199.8) | – | |
| Послеродовой | 649 | 42.3 (39.2–45.7) | 509 | 33.1 (30.4–36.1) | 135 | 27.4 (23.1–32.4) | 218 | 17.5 (15.3–20.0) |
| Ранний послеродовой период | 584 | 75.4 (69.6–81.8) | 460 | 59.3 (54.1–65.0) | 177 | 46.8 (39.1–56.1) | 199 | 30.4 (26.4–35.0) |
| Поздний послеродовой период | 65 | 8.5 (7.0–10.9) | 49 | 6.4 (4.9–8.5) | 18 | 7.3 (4.6–11.6) | 319 | 3.2 (1.9–5.0) |
| Коэффициенты заболеваемости (IRR) первой ВТЭ во время беременности и в послеродовом периоде | ||||||||
| Шведские данные А | Шведские данные B | Английские данные | Датские данные | |||||
| Период времени | ВНД* (95% ДИ) | ВНД* (95% ДИ) | ВНД (95% ДИ)† | ВНД (95% ДИ)† | ||||
| Вне беременности | Ссылка (т. е. 1,00) | |||||||
| Дородовый период | 5.08 (4.66–5.54) | 3.80 (3.44–4.19) | 3.10 (2.63–3.66) | 2.95 (2.68–3.25) | ||||
| Триместр 1 | 3.42 (2.95–3.98) | 3.04 (2.58–3.56) | 1.46 (0.96–2.20) | 1.12 (0.86–1.45) | ||||
| Триместр 2 | 4.31 (3.78–4.93) | 3.01 (2.56–3.53) | 1.82 (1.27–2.62) | 1.58 (1.24–1.99) | ||||
| Триместр 3 | 7.14 (6.43–7.94) | 5.12 (4.53–5.80) | 5.69 (4.66–6.95) | 5.48 (4.89–6.12) | ||||
| Вокруг доставки | 37.5 (30.9–44.45) | 27.97 (22.24–35.17) | 44.5 (31.68–62.54) | – | ||||
| Послеродовой | 10.21 (9.27–11.25) | 8.72 (7.83–9.70) | 8.54 (7.16–10.19) | 4.85 (4.21–5.57) | ||||
| Ранний послеродовой период | 19.27 (16.53–20.21) | 15.62 (14.00–17.45) | 14.61 (12.10–17.67) | 8.44 (7.27–9.75) | ||||
| Поздний послеродовой период | 2.06 (1.60–2.64) | 1.69 (1.26–2.25) | 2.29 (1.44–3.65) | 0.89 (0.53–1.39) | ||||
| Примечания: шведские данные A = использование любого кода для VTE независимо от подтверждения. Шведские данные B = использование только ВТЭ, подтвержденного алгоритмом. Ранний послеродовой период = первые 6 недель после родов. Поздний послеродовой период = более 6 недель после родов. * = Скорректировано с учетом возраста и календарного года. † = Нескорректированный коэффициент, рассчитанный на основе предоставленных данных. Источник: [88] | ||||||||
Иммунная система
[ редактировать ]Влияние эстрогена на иммунную систему в целом описывается как благоприятствующее Th2 , а не подавляющее, как в случае с действием мужского полового гормона - тестостерона. [89] Действительно, женщины лучше реагируют на вакцины , инфекции и, как правило, с меньшей вероятностью заболевают раком . Платой за это является то, что у них более вероятно развитие аутоиммунного заболевания . [90] Сдвиг Th2 проявляется в снижении клеточного иммунитета, а увеличение гуморального иммунитета ( выработки антител ) сдвигает его с клеточного на гуморальный за счет подавления клеточно-опосредованного иммунитета и усиления иммунного ответа Th2 за счет стимуляции продукции IL-4 и дифференцировки Th2. [89] [91] Иммунные реакции типов 1 и 17 подавляются, вероятно, по крайней мере частично, из-за IL-4 , который ингибирует Th1. Влияние эстрогена на различные типы клеток иммунных клеток соответствует его смещению Th2. Активность базофилов , эозинофилов М2 , макрофагов повышается, тогда как активность NK-клеток снижается. Обычные дендритные клетки склонны к Th2 под влиянием эстрогена, тогда как плазмоцитоидные дендритные клетки, ключевые игроки противовирусной защиты, увеличивают секрецию IFN-g . [91] Эстроген также влияет на В-клетки, увеличивая их выживаемость, пролиферацию, дифференцировку и функцию, что соответствует более высокому количеству антител и В-клеток, обычно выявляемому у женщин. [92]
На молекулярном уровне эстроген вызывает вышеупомянутые эффекты на клетку, воздействуя на внутриклеточные рецепторы , называемые ER α и ER β, которые при лигировании образуют либо гомо-, либо гетеродимеры. Генетические и негенетические мишени рецепторов различаются у гомо- и гетеродимеров. [93] Лигирование этих рецепторов позволяет им перемещаться в ядро и действовать как факторы транскрипции либо путем связывания элементов ответа на эстроген (ERE) с ДНК , либо связывания ДНК вместе с другими факторами транскрипции, например, Nf-kB или AP-1 , оба из которых приводят к образованию РНК. рекрутирование полимеразы и дальнейшая ремоделация хроматина . [93] Также был документирован нетранскрипционный ответ на стимуляцию эстрогеном (так называемый мембранно-инициируемый сигнал стероидов, MISS). Этот путь стимулирует пути ERK и PI3K/AKT, которые, как известно, увеличивают клеточную пролиферацию и влияют на ремоделацию хроматина. [93]
Сопутствующие условия
[ редактировать ]Исследователи связывают эстрогены с различными эстроген-зависимыми состояниями , такими как ER-положительный рак молочной железы , а также с рядом генетических состояний, включающих передачу сигналов или метаболизм эстрогена, таких как синдром нечувствительности к эстрогену , дефицит ароматазы и синдром избытка ароматазы .
Высокий уровень эстрогена может усиливать реакцию гормонов стресса в стрессовых ситуациях. [94]
Биохимия
[ редактировать ]Биосинтез
[ редактировать ]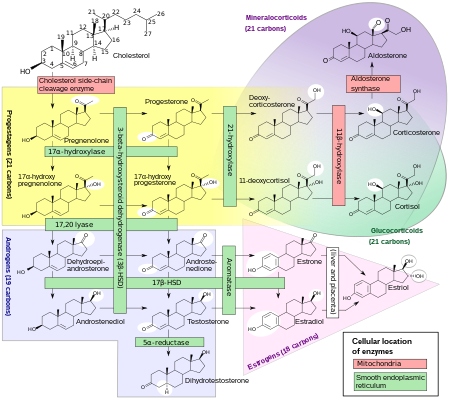
Эстрогены у женщин вырабатываются в основном яичниками , а во время беременности – плацентой . [96] Фолликулостимулирующий гормон (ФСГ) стимулирует выработку эстрогенов гранулезными клетками и фолликулов яичников желтых тел . Некоторые эстрогены также производятся в меньших количествах другими тканями, такими как печень , поджелудочная железа , кости , надпочечники , кожа , мозг , жировая ткань . [97] и грудь . [98] Эти вторичные источники эстрогенов особенно важны для женщин в постменопаузе. [99] Путь биосинтеза эстрогенов во внегонадных тканях различен. Эти ткани не способны синтезировать стероиды C19 и поэтому зависят от поставок C19 из других тканей. [99] и уровень ароматазы. [100]
У женщин синтез эстрогенов начинается в клетках внутренней теки яичника путем синтеза андростендиона из холестерина . Андростендион представляет собой вещество со слабой андрогенной активностью, которое служит преимущественно предшественником более мощных андрогенов, таких как тестостерон, а также эстроген. Это соединение проникает через базальную мембрану в окружающие гранулезные клетки, где оно превращается либо сразу в эстрон, либо в тестостерон, а затем на дополнительном этапе в эстрадиол. Превращение андростендиона в тестостерон катализируется 17β-гидроксистероиддегидрогеназой (17β-HSD), тогда как превращение андростендиона и тестостерона в эстрон и эстрадиол соответственно катализируется ароматазой, ферментами, которые экспрессируются в гранулезных клетках. Напротив, в гранулезных клетках отсутствует 17α-гидроксилаза и 17,20-лиаза , тогда как клетки теки экспрессируют эти ферменты и 17β-HSD, но лишены ароматазы. Следовательно, как гранулезные, так и тека-клетки необходимы для производства эстрогена в яичниках.
Уровни эстрогена варьируются в течение менструального цикла , причем самые высокие уровни достигаются ближе к концу фолликулярной фазы, непосредственно перед овуляцией .
Обратите внимание, что у мужчин эстроген также вырабатывается клетками Сертоли , когда ФСГ связывается с их рецепторами ФСГ.
Распределение
[ редактировать ]Эстрогены представляют собой белки плазмы, связанные с альбумином и/или глобулином, связывающим половые гормоны, в кровотоке.
Метаболизм
[ редактировать ]Эстрогены метаболизируются посредством гидроксилирования ферментами цитохрома P450 , такими как CYP1A1 и CYP3A4, а также посредством конъюгации с помощью сульфотрансфераз эстрогена ( сульфатирование ) и УДФ-глюкуронилтрансфераз ( глюкуронидация ). Кроме того, эстрадиол дегидрируется в 17β-гидроксистероиддегидрогеназой гораздо менее мощный эстроген эстрон. Эти реакции возникают преимущественно в печени , но также и в других тканях .
Метаболизм эстрогенов у человека |

Экскреция
[ редактировать ]Эстрогены инактивируются преимущественно почками и печенью и выводятся через желудочно-кишечный тракт. [101] в виде конъюгатов обнаруживается в кале , желчи и моче . [102]
Медицинское использование
[ редактировать ]Эстрогены используются в качестве лекарственных средств , главным образом в гормональной контрацепции , заместительной гормональной терапии , [103] и для лечения гендерной дисфории у трансгендерных женщин и других трансженщин в рамках феминизирующей гормональной терапии. [104]
Химия
[ редактировать ]Эстрогенные стероидные гормоны представляют собой эстрановые стероиды.
История
[ редактировать ]В 1929 году Адольф Бутенандт и Эдвард Адельберт Дойзи независимо друг от друга выделили и очистили эстрон, первый открытый эстроген. [105] Затем в 1930 и 1933 годах были открыты эстриол и эстрадиол соответственно. Вскоре после их открытия эстрогены, как природные, так и синтетические, стали использоваться в медицинских целях. Примеры включают глюкуронид эстриола ( Эмменин , Прогинон ), бензоат эстрадиола , конъюгированные эстрогены ( Премарин ), диэтилстильбэстрол и этинилэстрадиол .
Слово эстроген происходит от древнегреческого . Оно образовано от слова «эстрос». [106] (периодическое состояние половой активности самок млекопитающих) и генос (генерирующий). [106] Впервые он был опубликован в начале 1920-х годов и назывался «эстрин». [107] С годами американский английский адаптировал написание слова «эстроген» в соответствии с его фонетическим произношением.
Общество и культура
[ редактировать ]Этимология
[ редактировать ]Название эстроген происходит от греческого οἶστρος ( oîstros ), что буквально означает «воодушевление» или «вдохновение», а в переносном смысле — сексуальная страсть или желание. [108] и суффикс -gen , означающий «производитель».
Среда
[ редактировать ]выявлен ряд синтетических и природных веществ, обладающих эстрогенной активностью В окружающей среде и отнесенных к ксеноэстрогенам . [109]
- Синтетические вещества, такие как бисфенол А, а также металлоэстрогены (например, кадмий ).
- Растительные продукты с эстрогенной активностью называются фитоэстрогенами (например, куместрол , даидзеин , генистеин , мироэстрол ).
- Те, которые производятся грибами, известны как микоэстрогены (например, зеараленон ).
Эстрогены относятся к широкому спектру соединений, нарушающих работу эндокринной системы (EDC), поскольку они обладают высокой эстрогенной активностью. Когда EDC попадает в окружающую среду, он может вызвать мужскую репродуктивную дисфункцию у диких животных и людей. [12] [13] Эстроген, выделяемый сельскохозяйственными животными, попадает в системы пресной воды. [110] [111] В период прорастания и размножения рыбы подвергаются воздействию низкого уровня эстрогена, который может вызвать репродуктивную дисфункцию у самцов рыб. [112] [113]
Косметика
[ редактировать ]Некоторые шампуни для волос , представленные на рынке, содержат эстрогены и экстракты плаценты; другие содержат фитоэстрогены . В 1998 году были сообщения о случаях, когда у четырех афроамериканских девочек препубертатного возраста после воздействия этих шампуней появилась грудь. [114] В 1993 году FDA установило, что не все продаваемые без рецепта гормоносодержащие лекарственные средства местного применения для человека в целом признаны безопасными и эффективными и имеют неправильную торговую марку. Сопровождающее предлагаемое правило касается косметики и делает вывод, что любое использование натуральных эстрогенов в косметическом продукте делает этот продукт неутвержденным новым лекарственным средством и что любое косметическое средство, использующее термин «гормон» в тексте своей маркировки или в описании ингредиентов, подразумевает заявление о лекарственном препарате, подвергающее такой продукт регулятивным мерам. [115]
Продукты, в которых утверждается, что они содержат экстракт плаценты, не только считаются лекарствами с неправильной торговой маркой, но и могут считаться косметическими средствами с неправильной торговой маркой, если экстракт был приготовлен из плаценты, из которой были удалены гормоны и другие биологически активные вещества, и экстрагированное вещество состоит в основном из белка. . FDA рекомендует идентифицировать это вещество под названием, отличным от «экстракт плаценты», и более точно описывать его состав, поскольку у потребителей название «экстракт плаценты» ассоциируется с терапевтическим использованием некоторой биологической активности. [115]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Хьютер С.Э., Макканс К.Л. (2019). Понимание патофизиологии . Elsevier Науки о здоровье. п. 767. ИСБН 978-0-32-367281-8 .
Эстроген — это общий термин для любого из трех аналогичных гормонов, полученных из холестерина: эстрадиола, эстрона и эстриола.
- ^ Сатоскар Р.С., Реге Н., Бхандаркар С.Д. (2017). Фармакология и фармакотерапия . Elsevier Науки о здоровье. п. 943. ИСБН 978-8-13-124941-3 .
Природные эстрогены – это стероиды. Однако типичную эстрогенную активность проявляют и химические вещества, не являющиеся стероидами. Следовательно, термин «эстроген» используется как общий термин для описания всех соединений, обладающих эстрогенной активностью.
- ^ Дельгадо Б.Дж., Лопес-Охеда В. (20 декабря 2021 г.). «Эстроген» . StatPearls [Интернет] . Издательство StatPearls. PMID 30855848 .
Эстроген – это стероидный гормон, связанный с женскими репродуктивными органами и отвечающий за развитие женских половых признаков. Эстроген часто называют эстроном, эстрадиолом и эстриолом. ... Синтетический эстроген также доступен для клинического использования и предназначен для увеличения абсорбции и эффективности за счет изменения химической структуры эстрогена для местного или перорального применения. Синтетические стероидные эстрогены включают этинилэстрадиол, валерат эстрадиола, эстропипат, конъюгат эстерифицированного эстрогена и хинестрол.
- ^ Райан К.Дж. (август 1982 г.). «Биохимия ароматазы: значение для женской репродуктивной физиологии». Исследования рака . 42 (8 дополнений): 3342–3344 с. ПМИД 7083198 .
- ^ Мешулам Р., Брюггемайер Р.В., Денлингер Д.Л. (сентябрь 2005 г.). «Эстрогены у насекомых». Клеточные и молекулярные науки о жизни . 40 (9): 942–944. дои : 10.1007/BF01946450 . S2CID 31950471 .
- ^ Burger HG (апрель 2002 г.). «Продукция андрогенов у женщин» . Фертильность и бесплодие . 77 (Приложение 4): S3–S5. дои : 10.1016/S0015-0282(02)02985-0 . ПМИД 12007895 .
- ^ Ломбарди Г., Заррилли С., Колао А., Паэсано Л., Ди Сомма С., Росси Ф. и др. (июнь 2001 г.). «Эстрогены и здоровье мужчин». Молекулярная и клеточная эндокринология . 178 (1–2): 51–55. дои : 10.1016/S0303-7207(01)00420-8 . ПМИД 11403894 . S2CID 36834775 .
- ^ Уайтхед С.А., Насси С. (2001). Эндокринология: комплексный подход . Оксфорд: BIOS: Тейлор и Фрэнсис. ISBN 978-1-85996-252-7 . ПМИД 20821847 .
- ^ Солтысик К., Чекай П. (апрель 2013 г.). «Мембранные рецепторы эстрогена – альтернативный ли путь действия эстрогенов?». Журнал физиологии и фармакологии . 64 (2): 129–142. ПМИД 23756388 .
- ^ Мицевич П.Е., Келли М.Дж. (2012). «Мембранная регуляция функции гипоталамуса эстрогеновыми рецепторами» . Нейроэндокринология . 96 (2): 103–110. дои : 10.1159/000338400 . ПМЦ 3496782 . ПМИД 22538318 .
- ^ Просниц Э.Р., Артерберн Дж.Б., Склар Л.А. (февраль 2007 г.). «GPR30: рецептор эстрогена, связанный с белком AG» . Молекулярная и клеточная эндокринология . 265–266: 138–142. дои : 10.1016/j.mce.2006.12.010 . ПМЦ 1847610 . ПМИД 17222505 .
- ^ Перейти обратно: а б Ван С., Хуан В., Фан Г., Чжан Ю., Цяо Х. (2008). «Анализ остатков стероидных эстрогенов в образцах продуктов питания и окружающей среды». Международный журнал экологической аналитической химии . 88 (1): 1–25. Бибкод : 2008IJEAC..88....1W . дои : 10.1080/03067310701597293 . S2CID 93975613 .
- ^ Перейти обратно: а б Корач К.Д. (1998). Репродуктивная и онтотоксикология . Нью-Йорк: Марсель Деккер. ISBN 0-585-15807-Х . OCLC 44957536 .
- ^ А. Лабхарт (6 декабря 2012 г.). Клиническая эндокринология: теория и практика . Springer Science & Business Media. стр. 548–. ISBN 978-3-642-96158-8 .
- ^ Бейкер М.Э. (март 2013 г.). «Что такое физиологические эстрогены?». Стероиды . 78 (3): 337–340. doi : 10.1016/j.steroids.2012.12.011 . ПМИД 23313336 . S2CID 11803629 .
- ^ Миллер К.К., Аль-Райян Н., Иванова М.М., Маттингли К.А., Рипп С.Л., Клинге С.М. и др. (январь 2013 г.). «Метаболиты DHEA активируют рецепторы эстрогена альфа и бета» . Стероиды . 78 (1): 15–25. doi : 10.1016/j.steroids.2012.10.002 . ПМЦ 3529809 . ПМИД 23123738 .
- ^ Бхавнани Б.Р., Нискер Дж.А., Мартин Дж., Алетеби Ф., Уотсон Л., Милн Дж.К. (2000). «Сравнение фармакокинетики препарата конъюгированного лошадиного эстрогена (премарин) и синтетической смеси эстрогенов (CES) у женщин в постменопаузе». Журнал Общества гинекологических исследований . 7 (3): 175–183. дои : 10.1016/s1071-5576(00)00049-6 . ПМИД 10865186 .
- ^ Хэггстрем М (2014). «Референтные диапазоны эстрадиола, прогестерона, лютеинизирующего гормона и фолликулостимулирующего гормона во время менструального цикла» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.001 . ISSN 2002-4436 .
- ^ Лин С.И., Стрём А., Вега В.Б., Конг С.Л., Йео А.Л., Томсен Дж.С. и др. (2004). «Открытие генов-мишеней альфа-рецептора эстрогена и элементов ответа в клетках опухоли молочной железы» . Геномная биология . 5 (9): С66. дои : 10.1186/gb-2004-5-9-r66 . ПМК 522873 . ПМИД 15345050 .
- ^ Дараби М., Ани М., Панджепур М., Раббани М., Мовахедиан А., Зареан Э. (2011). «Влияние полиморфизма рецептора эстрогена β A1730G на реакцию экспрессии гена ABCA1 на заместительную гормональную терапию в постменопаузе». Генетическое тестирование и молекулярные биомаркеры . 15 (1–2): 11–15. дои : 10.1089/gtmb.2010.0106 . ПМИД 21117950 .
- ^ Лауэрс Дж., Шински Д. (2004). Консультирование кормящей матери: Руководство консультанта по грудному вскармливанию . Джонс и Бартлетт Лиринг, ООО. п. 93. ИСБН 978-0-7637-2765-9 . Проверено 12 октября 2023 г.
- ^ Ралофф Дж. (6 декабря 1997 г.). «Science News Online (6.12.97): Возникающее мужественное альтер-эго эстрогена» . Новости науки . Проверено 4 марта 2008 г.
- ^ Хесс Р.А., Буник Д., Ли К.Х., Бар Дж., Тейлор Дж.А., Корах К.С. и др. (декабрь 1997 г.). «Роль эстрогенов в мужской репродуктивной системе» . Природа . 390 (6659): 509–512. Бибкод : 1997Natur.390..509H . дои : 10.1038/37352 . ПМЦ 5719867 . ПМИД 9393999 .
- ^ «Эстроген связан с количеством сперматозоидов и мужской фертильностью» . Научный блог. Архивировано из оригинала 7 мая 2007 года . Проверено 4 марта 2008 г.
- ^ Хилл Р.А., Помполо С., Джонс М.Э., Симпсон Э.Р., Бун У.К. (декабрь 2004 г.). «Дефицит эстрогена приводит к апоптозу дофаминергических нейронов в медиальной преоптической области и дугообразном ядре самцов мышей». Молекулярная и клеточная нейронауки . 27 (4): 466–476. дои : 10.1016/j.mcn.2004.04.012 . ПМИД 15555924 . S2CID 25280077 .
- ^ Чиди-Огболу Н., Баар К. (2018). «Влияние эстрогена на работу скелетно-мышечной системы и риск травм» . Границы в физиологии . 9 : 1834. doi : 10.3389/fphys.2018.01834 . ПМК 6341375 . ПМИД 30697162 .
- ^ Лоу Д.А., Балтгалвис К.А., Грайзинг С.М. (апрель 2010 г.). «Механизмы благотворного влияния эстрогена на мышечную силу у женщин» . Обзоры физических упражнений и спортивных наук . 38 (2): 61–67. дои : 10.1097/JES.0b013e3181d496bc . ПМЦ 2873087 . ПМИД 20335737 .
- ^ Макс С.Р. (декабрь 1984 г.). «Синергия андрогенов и эстрогенов в мышце, поднимающей задний проход крысы: глюкозо-6-фосфатдегидрогеназа». Молекулярная и клеточная эндокринология . 38 (2–3): 103–107. дои : 10.1016/0303-7207(84)90108-4 . ПМИД 6510548 . S2CID 24198956 .
- ^ Кут Р.В., Амелинк Г.Дж., Бланкенштейн М.А., Бэр П.Р. (1991). «Тамоксифен и эстроген защищают мышцы крысы от физиологических повреждений». Журнал биохимии стероидов и молекулярной биологии . 40 (4–6): 689–695. дои : 10.1016/0960-0760(91)90292-д . ПМИД 1958566 . S2CID 44446541 .
- ^ Хайзлип К.М., Харрисон BC, Лейнванд Л.А. (январь 2015 г.). «Половые различия в кинетике скелетных мышц и составе волокон» . Физиология . 30 (1): 30–39. дои : 10.1152/физиол.00024.2014 . ПМЦ 4285578 . ПМИД 25559153 . «Добавка эстрогена увеличивает процентное содержание типа III в подошвенной мышце обратно до 42%. (70)»
- ^ Фрэнк А.П., де Соуза Сантос Р., Палмер Б.Ф., Клегг DJ (октябрь 2019 г.). «Детерминанты распределения жира в организме человека могут дать представление о рисках для здоровья, связанных с ожирением» . Журнал исследований липидов . 60 (10): 1710–1719. дои : 10.1194/jlr.R086975 . ПМК 6795075 . ПМИД 30097511 .
- ^ Браун Л.М., Гент Л., Дэвис К., Клегг DJ (сентябрь 2010 г.). «Метаболическое влияние половых гормонов на ожирение» . Исследования мозга . 1350 : 77–85. дои : 10.1016/j.brainres.2010.04.056 . ПМЦ 2924463 . ПМИД 20441773 .
- ^ Янссен И., Пауэлл Л.Х., Казлаускайте Р., Дуган С.А. (март 2010 г.). «Тестостерон и висцеральный жир у женщин среднего возраста: исследование структуры жира в исследовании женского здоровья по всей стране (SWAN)» . Ожирение . 18 (3): 604–610. дои : 10.1038/oby.2009.251 . ПМЦ 2866448 . ПМИД 19696765 .
- ^ Рубинов КБ (2017). «Эстрогены и регулирование массы тела у мужчин». Половые и гендерные факторы, влияющие на метаболический гомеостаз, диабет и ожирение . Достижения экспериментальной медицины и биологии. Том. 1043. Спрингер. стр. 285–313. дои : 10.1007/978-3-319-70178-3_14 . ISBN 978-3-319-70177-6 . ПМЦ 5835337 . ПМИД 29224100 .
- ^ Перейти обратно: а б Стокар Дж., Гурт И., Коэн-Кфир Э., Якубовский О., Халлак Н., Беньямини Х. и др. (июнь 2022 г.). «Печеночный адропин регулируется эстрогеном и способствует неблагоприятным метаболическим фенотипам у мышей с удаленными яичниками» . Молекулярный метаболизм . 60 : 101482. doi : 10.1016/j.molmet.2022.101482 . ПМК 9044006 . ПМИД 35364299 .
- ^ Фрайш П. «Причины, по которым ваше лицо выглядит опухшим» . ВебМД .
- ^ Стахенфельд Н.С. (июль 2008 г.). «Влияние половых гормонов на регуляцию жидкости в организме» . Обзоры физических упражнений и спортивных наук . 36 (3): 152–159. дои : 10.1097/JES.0b013e31817be928 . ПМЦ 2849969 . ПМИД 18580296 .
- ^ Павлина В (2023). Гистология: текст и атлас: с коррелирующей клеточной и молекулярной биологией . Уолтерс Клювер Здоровье. п. 1481. ИСБН 978-1-9751-8152-9 . Проверено 12 октября 2023 г.
- ^ Гринберг Дж., Брюсс С., Освальт С. (2014). «Зачатие, беременность и роды» . Исследование аспектов человеческой сексуальности . Джонс и Бартлетт Обучение. п. 248. ИСБН 978-1-4496-4851-0 . Проверено 12 октября 2023 г.
- ^ «Исследователи обнаруживают генетические причины повышенного риска развития меланомы у мужчин» . ScienceDaily .
- ^ Эрнандо Б., Ибаррола-Вильява М., Фернандес Л.П., Пенья-Чиле М., Льорка-Карденоса М., Ольтра СС и др. (18 марта 2016 г.). «Полоспецифичные генетические эффекты, связанные с пигментацией, чувствительностью к солнечному свету и меланомой в популяции испанского происхождения» . Биология половых различий . 7 (1): 17. дои : 10.1186/s13293-016-0070-1 . ПМЦ 4797181 . ПМИД 26998216 . «Результаты этого исследования показывают, что действительно существуют генетические эффекты, специфичные для пола, в пигментации человека, с более сильным эффектом для более темной пигментации у женщин по сравнению с мужчинами. Вероятной причиной может быть дифференциальная экспрессия меланогенных генов у женщин из-за более высоких уровней эстрогена. Эти генетические эффекты, специфичные для пола, могут помочь объяснить наличие более темной пигментации глаз и кожи у женщин, а также хорошо известный более высокий риск развития меланомы у мужчин».
- ^ Массаро Д., Массаро Г.Д. (декабрь 2004 г.). «Эстроген регулирует образование, потерю и регенерацию легочных альвеол у мышей» (PDF) . Американский журнал физиологии. Клеточная и молекулярная физиология легких . 287 (6): Л1154–Л1159. дои : 10.1152/ajplung.00228.2004 . ПМИД 15298854 . S2CID 24642944 . Архивировано из оригинала (PDF) 25 февраля 2019 года.
- ^ Кристенсен А., Бентли Дж.Э., Кабрера Р., Ортега Х.Х., Перфито Н., Ву Т.Дж. и др. (июль 2012 г.). «Гормональная регуляция женской репродукции» . Гормональные и метаболические исследования . 44 (8): 587–591. дои : 10.1055/s-0032-1306301 . ПМЦ 3647363 . ПМИД 22438212 .
- ^ Ханда Р.Дж., Огава С., Ван Дж.М., Хербисон А.Е. (январь 2012 г.). «Роль эстрогенового рецептора β в функции мозга взрослого человека» . Журнал нейроэндокринологии . 24 (1): 160–173. дои : 10.1111/j.1365-2826.2011.02206.x . ПМЦ 3348521 . ПМИД 21851428 .
- ^ Коу Л.М., Пфафф Д.В. (май 1998 г.). «Картирование путей нейронной и сигнальной трансдукции лордоза в поисках действия эстрогена на центральную нервную систему». Поведенческие исследования мозга . 92 (2): 169–180. дои : 10.1016/S0166-4328(97)00189-7 . ПМИД 9638959 . S2CID 28276218 .
- ^ Уорнок Дж.К., Суонсон С.Г., Борель Р.В., Зипфель Л.М., Бреннан Дж.Дж. (2005). «Комбинированные этерифицированные эстрогены и метилтестостерон по сравнению с одними этерифицированными эстрогенами при лечении потери сексуального интереса у женщин в хирургической менопаузе». Менопауза . 12 (4): 374–384. doi : 10.1097/01.GME.0000153933.50860.FD . ПМИД 16037752 . S2CID 24557071 .
- ^ Хейман-младший, Рупп Х., Янссен Э., Ньюхаус С.К., Брауэр М., Лаан Э. (май 2011 г.). «Сексуальное желание, сексуальное возбуждение и гормональные различия у женщин в США и Нидерландах в пременопаузе с низким сексуальным желанием и без него». Гормоны и поведение . 59 (5): 772–779. дои : 10.1016/j.yhbeh.2011.03.013 . ПМИД 21514299 . S2CID 20807391 .
- ^ Брискен С., О'Мэлли Б (декабрь 2010 г.). «Действие гормонов в молочной железе» . Перспективы Колд-Спринг-Харбор в биологии . 2 (12): а003178. doi : 10.1101/cshperspect.a003178 . ПМЦ 2982168 . ПМИД 20739412 .
- ^ Кляйнберг Д.Л. (февраль 1998 г.). «Роль IGF-I в нормальном развитии молочной железы». Исследование и лечение рака молочной железы . 47 (3): 201–208. дои : 10.1023/а:1005998832636 . ПМИД 9516076 . S2CID 30440069 .
- ^ Перейти обратно: а б с Джонсон Л.Р. (2003). Основная медицинская физиология . Академическая пресса. п. 770. ИСБН 978-0-12-387584-6 .
- ^ Перейти обратно: а б с Норман А.В., Генри Х.Л. (30 июля 2014 г.). Гормоны . Академическая пресса. п. 311. ИСБН 978-0-08-091906-5 .
- ^ Перейти обратно: а б Коад Дж., Данстолл М. (2011). Анатомия и физиология для акушерок, с онлайн-доступом Pageburst, 3: Анатомия и физиология для акушерок . Elsevier Науки о здоровье. п. 413. ИСБН 978-0-7020-3489-3 .
- ^ Хаслам С.З., Осуч-младший (1 января 2006 г.). Гормоны и рак молочной железы у женщин в постменопаузе . ИОС Пресс. п. 69. ИСБН 978-1-58603-653-9 .
- ^ Зильбернагль С., Деспопулос А (1 января 2011 г.). Цветной атлас физиологии . Тиме. стр. 305–. ISBN 978-3-13-149521-1 .
- ^ Фадем Б (2007). Высокопроизводительный комплексный обзор USMLE, шаг 1 . Липпинкотт Уильямс и Уилкинс. стр. 445–. ISBN 978-0-7817-7427-7 .
- ^ Блэкберн С (14 апреля 2014 г.). Физиология матери, плода и новорожденного . Elsevier Науки о здоровье. стр. 146–. ISBN 978-0-323-29296-2 .
- ^ Штраус Дж. Ф., Барбьери Р. Л. (13 сентября 2013 г.). Репродуктивная эндокринология Йена и Яффе . Elsevier Науки о здоровье. стр. 236–. ISBN 978-1-4557-2758-2 .
- ^ Уилсон CB, Низет В., Мальдонадо Й., Ремингтон Дж.С., Кляйн Дж.О. (24 февраля 2015 г.). Инфекционные болезни плода и новорожденного Ремингтона и Кляйна . Elsevier Науки о здоровье. стр. 190–. ISBN 978-0-323-24147-2 .
- ^ Сарате С., Стевнснер Т., Гредилья Р. (2017). «Роль эстрогена и других половых гормонов в старении мозга. Нейропротекция и восстановление ДНК» . Границы стареющей неврологии . 9 : 430. дои : 10.3389/fnagi.2017.00430 . ПМЦ 5743731 . ПМИД 29311911 .
- ^ Шервин Б.Б. (февраль 2012 г.). «Эстроген и когнитивное функционирование у женщин: уроки, которые мы извлекли» . Поведенческая нейронаука . 126 (1): 123–127. дои : 10.1037/a0025539 . ПМЦ 4838456 . ПМИД 22004260 .
- ^ Хара Ю., Уотерс Э.М., МакИвен Б.С., Моррисон Дж.Х. (июль 2015 г.). «Влияние эстрогена на когнитивное и синаптическое здоровье на протяжении всей жизни» . Физиологические обзоры . 95 (3): 785–807. doi : 10.1152/physrev.00036.2014 . ПМЦ 4491541 . ПМИД 26109339 .
- ^ Король Д.Л., Пизани С.Л. (август 2015 г.). «Эстрогены и познание: друзья или враги?: Оценка противоположного воздействия эстрогенов на обучение и память» . Гормоны и поведение . 74 : 105–115. дои : 10.1016/j.yhbeh.2015.06.017 . ПМЦ 4573330 . ПМИД 26149525 .
- ^ Ау А, Фехер А, Макфи Л, Джесса А, О С, Эйнштейн Г (январь 2016 г.). «Эстрогены, воспаление и познание» . Границы нейроэндокринологии . 40 : 87–100. doi : 10.1016/j.yfrne.2016.01.002 . ПМИД 26774208 .
- ^ Джейкобс Э., Д'Эспозито М. (апрель 2011 г.). «Эстроген формирует дофамин-зависимые когнитивные процессы: последствия для здоровья женщин» . Журнал неврологии . 31 (14): 5286–5293. doi : 10.1523/JNEUROSCI.6394-10.2011 . ПМК 3089976 . ПМИД 21471363 .
- ^ Колзато Л.С., Хоммель Б. (1 января 2014 г.). «Влияние эстрогена на когнитивные функции высшего порядка у женщин, не подвергающихся стрессу, может зависеть от индивидуальных различий в базовых уровнях дофамина» . Границы в неврологии . 8 : 65. дои : 10.3389/fnins.2014.00065 . ПМЦ 3985021 . ПМИД 24778605 .
- ^ Хогерворст Э (март 2013 г.). «Эстроген и мозг: улучшает ли лечение эстрогенами когнитивные функции?». Менопауза Интернэшнл . 19 (1): 6–19. дои : 10.1177/1754045312473873 . ПМИД 27951525 . S2CID 10122688 .
- ^ Дума С.Л., Муж С., О'Доннелл М.Э., Барвин Б.Н., Вуденд А.К. (2005). «Расстройства настроения, связанные с эстрогеном: факторы репродуктивного жизненного цикла». АНС. Достижения в области сестринского дела . 28 (4): 364–375. дои : 10.1097/00012272-200510000-00008 . ПМИД 16292022 . S2CID 9172877 .
- ^ Остерлунд М.К., Витт М.Р., Густафссон Дж.А. (декабрь 2005 г.). «Действие эстрогена на настроение и нейродегенеративные расстройства: эстрогенные соединения с селективными свойствами - новое поколение терапевтических средств». Эндокринный . 28 (3): 235–242. дои : 10.1385/ЭНДО:28:3:235 . ПМИД 16388113 . S2CID 8205014 .
- ^ Ласюк Г.Ц., Хегадорен К.М. (октябрь 2007 г.). «Влияние эстрадиола на центральные серотонинергические системы и его связь с настроением у женщин». Биологические исследования для медсестер . 9 (2): 147–160. дои : 10.1177/1099800407305600 . ПМИД 17909167 . S2CID 37965502 .
- ^ Григориадис С., Симан М.В. (июнь 2002 г.). «Роль эстрогена при шизофрении: значение для практических рекомендаций по лечению шизофрении для женщин» . Канадский журнал психиатрии . 47 (5): 437–442. дои : 10.1177/070674370204700504 . ПМИД 12085678 .
- ^ «ПМДД/ПМС» . Центр психического здоровья женщин Массачусетской больницы общего профиля . Проверено 12 января 2019 г. .
- ^ Хилл Р.А., Макиннес К.Дж., Гонг Э.К., Джонс М.Э., Симпсон Э.Р., Бун У.К. (февраль 2007 г.). «У самцов мышей с дефицитом эстрогена развивается компульсивное поведение». Биологическая психиатрия . 61 (3): 359–366. doi : 10.1016/j.biopsych.2006.01.012 . ПМИД 16566897 . S2CID 22669945 .
- ^ Бенмансур С., Уивер Р.С., Бартон А.К., Адениджи О.С., Фрейзер А. (апрель 2012 г.). «Сравнение влияния эстрадиола и прогестерона на серотонинергическую функцию» . Биологическая психиатрия . 71 (7): 633–641. doi : 10.1016/j.biopsych.2011.11.023 . ПМК 3307822 . ПМИД 22225849 .
- ^ Берг С.Дж., Винн-Эдвардс К.Э. (июнь 2001 г.). «Изменения уровней тестостерона, кортизола и эстрадиола у мужчин, становящихся отцами». Труды клиники Мэйо . 76 (6): 582–592. дои : 10.4065/76.6.582 . ПМИД 11393496 .
- ^ Перейти обратно: а б с Цао X, Сюй П., Ойола М.Г., Ся Y, Ян X, Сайто К. и др. (октябрь 2014 г.). «Эстрогены стимулируют серотониновые нейроны, препятствуя перееданию у мышей» . Журнал клинических исследований . 124 (10): 4351–4362. дои : 10.1172/JCI74726 . ПМК 4191033 . ПМИД 25157819 .
- ^ Джимерсон, округ Колумбия, Лесем, доктор медицинских наук, Кэй У.Х., Хегг А.П., Брюэртон Т.Д. (сентябрь 1990 г.). «Расстройства пищевого поведения и депрессия: есть ли связь с серотонином?». Биологическая психиатрия . 28 (5): 443–454. дои : 10.1016/0006-3223(90)90412-у . ПМИД 2207221 . S2CID 31058047 .
- ^ Кламп К.Л., Кил П.К., Расин С.Е., Берт С.А., Берт А.С., Нил М. и др. (февраль 2013 г.). «Взаимодействующее воздействие эстрогена и прогестерона на изменения эмоционального питания в течение менструального цикла» . Журнал аномальной психологии . 122 (1): 131–137. дои : 10.1037/a0029524 . ПМК 3570621 . ПМИД 22889242 .
- ^ Перейти обратно: а б Эдлер С., Липсон С.Ф., Кил П.К. (январь 2007 г.). «Гормоны яичников и переедание при нервной булимии». Психологическая медицина . 37 (1): 131–141. дои : 10.1017/S0033291706008956 . ПМИД 17038206 . S2CID 36609028 .
- ^ Перейти обратно: а б Кламп К.Л., Расин С.Е., Хильдебрандт Б., Берт С.А., Нил М., Сиск К.Л. и др. (сентябрь 2014 г.). «Влияние гормонов яичников на нерегулируемое питание: сравнение ассоциаций у женщин с эпизодами переедания и без них» . Клиническая психологическая наука . 2 (4): 545–559. дои : 10.1177/2167702614521794 . ПМК 4203460 . ПМИД 25343062 .
- ^ Кламп К.Л., Кил П.К., Калберт К.М., Эдлер С. (декабрь 2008 г.). «Гормоны яичников и переедание: изучение связей в общественных образцах» . Психологическая медицина . 38 (12): 1749–1757. дои : 10.1017/S0033291708002997 . ПМЦ 2885896 . ПМИД 18307829 .
- ^ Лестер Н.А., Кил П.К., Липсон С.Ф. (январь 2003 г.). «Колебание симптомов при нервной булимии: связь с фазой менструального цикла и уровнем кортизола». Психологическая медицина . 33 (1): 51–60. дои : 10.1017/s0033291702006815 . ПМИД 12537036 . S2CID 21497515 .
- ^ Ву М.В., Маноли Д.С., Фрейзер Э.Дж., Коутс Дж.К., Толлкун Дж., Хонда С. и др. (октябрь 2009 г.). «Эстроген маскулинизирует нервные пути и половое поведение» . Клетка . 139 (1): 61–72. дои : 10.1016/j.cell.2009.07.036 . ПМЦ 2851224 . ПМИД 19804754 .
- ^ Рочира В., Карани С. (октябрь 2009 г.). «Дефицит ароматазы у мужчин: клиническая перспектива» . Обзоры природы. Эндокринология . 5 (10): 559–568. дои : 10.1038/nrendo.2009.176 . ПМИД 19707181 . S2CID 22116130 .
- ^ Уилсон Дж.Д. (сентябрь 2001 г.). «Андрогены, андрогенные рецепторы и мужское гендерно-ролевое поведение» (PDF) . Гормоны и поведение . 40 (2): 358–366. дои : 10.1006/hbeh.2001.1684 . ПМИД 11534997 . S2CID 20480423 . Архивировано из оригинала (PDF) 26 февраля 2019 года.
- ^ Баум М.Дж. (ноябрь 2006 г.). «Модели психосексуальной дифференциации млекопитающих: когда возможен «перенос» на человеческую ситуацию?». Гормоны и поведение . 50 (4): 579–588. дои : 10.1016/j.yhbeh.2006.06.003 . ПМИД 16876166 . S2CID 7465192 .
- ^ Розано ГМ, Панина Г (1999). «Эстрогены и сердце». Терапия . 54 (3): 381–385. ПМИД 10500455 .
- ^ Надкарни С., Купер Д., Бранкалеоне В., Бена С., Перретти М. (ноябрь 2011 г.). «Активация пути аннексина А1 лежит в основе защитного действия эстрогена на полиморфно-ядерные лейкоциты» . Атеросклероз, тромбоз и сосудистая биология . 31 (11): 2749–2759. дои : 10.1161/ATVBAHA.111.235176 . ПМЦ 3357483 . ПМИД 21836070 .
- ^ Абдул Султан А., Уэст Дж., Стефанссон О., Грейндж М.Дж., Тата Л.Дж., Флеминг К.М. и др. (ноябрь 2015 г.). «Определение венозной тромбоэмболии и измерение ее заболеваемости с использованием шведских реестров здравоохранения: общенациональное когортное исследование беременных» . БМЖ Опен . 5 (11): e008864. doi : 10.1136/bmjopen-2015-008864 . ПМЦ 4654387 . ПМИД 26560059 .
- ^ Перейти обратно: а б Фу Ю.З., Накагава С., Роудс Дж., Симмонс Л.В. (февраль 2017 г.). «Влияние половых гормонов на иммунную функцию: метаанализ» (PDF) . Биологические обзоры Кембриджского философского общества . 92 (1): 551–571. дои : 10.1111/brv.12243 . ПМИД 26800512 . S2CID 37931012 .
- ^ Танея В. (27 августа 2018 г.). «Половые гормоны определяют иммунный ответ» . Границы в иммунологии . 9 : 1931. дои : 10.3389/fimmu.2018.01931 . ПМК 6119719 . ПМИД 30210492 .
- ^ Перейти обратно: а б Ровед Дж., Вестердал Х., Хассельквист Д. (февраль 2017 г.). «Половые различия в иммунных реакциях: гормональные эффекты, антагонистический отбор и эволюционные последствия». Гормоны и поведение . 88 : 95–105. дои : 10.1016/j.yhbeh.2016.11.017 . ПМИД 27956226 . S2CID 9137227 .
- ^ Хан Д., Ансар Ахмед С. (6 января 2016 г.). «Иммунная система является естественной мишенью для действия эстрогена: противоположные эффекты эстрогена при двух прототипных аутоиммунных заболеваниях» . Границы в иммунологии . 6 : 635. дои : 10.3389/fimmu.2015.00635 . ПМК 4701921 . ПМИД 26779182 .
- ^ Перейти обратно: а б с Ковац С (апрель 2015 г.). «Эстрогеновые рецепторы регулируют врожденные иммунные клетки и сигнальные пути» . Клеточная иммунология . 294 (2): 63–69. дои : 10.1016/j.cellimm.2015.01.018 . ПМК 4380804 . ПМИД 25682174 .
- ^ Приор JC (2018). Сезон эстрогеновой бури: истории перименопаузы . Ванкувер, Британская Колумбия: CeMCOR (Центр исследований менструального цикла и овуляции). ISBN 9780973827521 . Проверено 24 июля 2021 г.
[...] высокий уровень эстрогена усиливает реакцию гормона стресса на стрессовые [...]
- ^ Хэггстрем М., Ричфилд Д. (2014). «Схема путей стероидогенеза человека» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.005 . ISSN 2002-4436 .
- ^ Мариб Э (2013). Анатомия и физиология . Бенджамин-Каммингс. п. 903. ИСБН 978-0-321-88760-3 .
- ^ Хемселл Д.Л., Гродин Дж.М., Бреннер П.Ф., Сиитери П.К., Макдональд ПК (март 1974 г.). «Плазменные предшественники эстрогена. II. Корреляция степени превращения плазменного андростендиона в эстрон с возрастом» . Журнал клинической эндокринологии и метаболизма . 38 (3): 476–479. doi : 10.1210/jcem-38-3-476 . ПМИД 4815174 .
- ^ Баракат Р., Окли О, Ким Х, Джин Дж, Ко СиДжей (сентябрь 2016 г.). «Внегонадные участки биосинтеза и функции эстрогенов» . Отчеты БМБ . 49 (9): 488–496. дои : 10.5483/BMBRep.2016.49.9.141 . ПМК 5227141 . ПМИД 27530684 .
- ^ Перейти обратно: а б Нельсон Л.Р., Булун С.Е. (сентябрь 2001 г.). «Производство и действие эстрогена». Журнал Американской академии дерматологии . 45 (3 дополнения): S116–S124. дои : 10.1067/mjd.2001.117432 . ПМИД 11511861 .
- ^ Лабри Ф., Беланжер А., Луу-Те В., Лабри С., Симард Дж., Кузан Л. и др. (1998). «ДГЭА и интракринное образование андрогенов и эстрогенов в периферических тканях-мишенях: его роль во время старения». Стероиды . 63 (5–6): 322–328. дои : 10.1016/S0039-128X(98)00007-5 . ПМИД 9618795 . S2CID 37344052 .
- ^ Шрейдер В.Е. (2012). «Яичник» . В: Тракслер А., Торн Г., Лабхарт А., Бюрги Х., Додсворт-Филлипс Дж., Констам Г., Курвуазье Б., Фишер Дж.А., Фрёш Э.Р., Гроб П. (ред.). Клиническая эндокринология: теория и практика . Шпрингер Берлин Гейдельберг. п. 530. ИСБН 978-3-642-96158-8 . Проверено 12 октября 2023 г.
- ^ Фуэнтес Н., Сильвейра П. (2019). «Механизмы передачи сигналов рецептора эстрогена». Достижения в области химии белков и структурной биологии . Том. 116. Эльзевир. стр. 135–170. дои : 10.1016/bs.apcsb.2019.01.001 . ISBN 9780128155615 . ISSN 1876-1623 . ПМК 6533072 . ПМИД 31036290 .
Физиологически метаболическое преобразование эстрогенов обеспечивает их выведение из организма через мочу, фекалии и/или желчь, а также выработку аналогов эстрогена, которые, как было показано, обладают антипролиферативным действием (Tsuriya et al., 2005).
- ^ Куль Х. (август 2005 г.). «Фармакология эстрогенов и прогестагенов: влияние различных путей введения». Климактерический . 8 (Приложение 1): 3–63. дои : 10.1080/13697130500148875 . ПМИД 16112947 . S2CID 24616324 .
- ^ Весп Л.М., Deutsch MB (март 2017 г.). «Варианты гормонального и хирургического лечения трансгендерных женщин и лиц трансженского спектра». Психиатрические клиники Северной Америки . 40 (1): 99–111. дои : 10.1016/j.psc.2016.10.006 . ПМИД 28159148 .
- ^ Тата-младший (июнь 2005 г.). «Сто лет гормонов» . Отчеты ЭМБО . 6 (6): 490–496. дои : 10.1038/sj.embor.7400444 . ПМЦ 1369102 . ПМИД 15940278 .
- ^ Перейти обратно: а б «Происхождение с точки зрения биомедицины: эстроген или эстроген» . Биоэтимология . Проверено 24 января 2018 г.
- ^ «Совет по фармации и химии». Журнал Американской медицинской ассоциации . 107 (15): 1221–3. 1936. дои : 10.1001/jama.1936.02770410043011 .
- ^ «Инструмент для изучения греческого слова: oistros» . Цифровая библиотека Персея . Проверено 28 декабря 2011 г.
- ^ Фанг Х., Тонг В., Ши Л.М., Блэр Р., Перкинс Р., Бранхам В. и др. (март 2001 г.). «Взаимосвязь структура-активность для большого разнообразного набора природных, синтетических и экологических эстрогенов». Химические исследования в токсикологии . 14 (3): 280–294. CiteSeerX 10.1.1.460.20 . дои : 10.1021/tx000208y . ПМИД 11258977 .
- ^ Уайз А., О'Брайен К., Вудрафф Т. (январь 2011 г.). «Вносят ли оральные контрацептивы значительный вклад в эстрогенность питьевой воды?». Экологические науки и технологии . 45 (1): 51–60. дои : 10.1021/es1014482 . ПМИД 20977246 .
- ^ Пич С. «Не вините таблетку | Последние новости» . Новости химии и техники . Проверено 22 апреля 2023 г.
- ^ Лини К.Э., Джоблинг С., Ширс Дж.А., Симпсон П., Тайлер Ч.Р. (октябрь 2005 г.). «Оценка чувствительности различных стадий жизни к половым нарушениям у плотвы (Rutilus rutilus), подвергшейся воздействию сточных вод очистных сооружений» . Перспективы гигиены окружающей среды . 113 (10): 1299–1307. дои : 10.1289/ehp.7921 . ПМЦ 1281270 . ПМИД 16203238 .
- ^ Джоблинг С., Уильямс Р., Джонсон А., Тейлор А., Гросс-Сорокин М., Нолан М. и др. (апрель 2006 г.). «Прогнозируемое воздействие стероидных эстрогенов в реках Великобритании коррелирует с широко распространенными сексуальными нарушениями в популяциях диких рыб» . Перспективы гигиены окружающей среды . 114 (Приложение 1): 32–39. дои : 10.1289/ehp.8050 . ПМЦ 1874167 . ПМИД 16818244 .
- ^ Сангхави Д.М. (17 октября 2006 г.). «Дошкольное половое созревание и поиск причин» . Нью-Йорк Таймс . Проверено 4 июня 2008 г.
- ^ Перейти обратно: а б FDA (февраль 1995 г.). «Продукты, содержащие эстрогенные гормоны, экстракт плаценты или витамины» . Руководство по проверкам производителей косметической продукции . Архивировано из оригинала 14 октября 2007 года . Проверено 24 октября 2006 г.
Внешние ссылки
[ редактировать ]- Насси и Уайтхед: Эндокринология, комплексный подход , Тейлор и Фрэнсис, 2001. Бесплатный онлайн-учебник.
- Фенолы
- Циклопентанолы
- Антигонадотропины
- Стране
- Эстрогены
- Фертильность
- Гепатотоксины
- Гормоны оси гипоталамус-гипофиз-гонады
- Гормоны гипоталамо-гипофизарно-пролактиновой оси
- Гормоны яичников
- Гормоны беременной женщины
- Гормоны яичка
- Женская эндокринная система человека
- Человеческие гормоны
- Женская репродуктивная система млекопитающих
- Релизеры пролактина
- Половые гормоны
- 1929 год в науке
- 1929 год в Германии.
- Стеролы
- Половое созревание