Дофамин
 Скелетная формула дофамина | |
 шарика и шарика, Модель молекулы дофамина как обнаружено в растворе. В твердом состоянии дофамин принимает Zwitterionic Form. [ 1 ] [ 2 ] | |
| Клинические данные | |
|---|---|
| Другие имена |
|
| Физиологические данные | |
| Исходные ткани | Субстанция nigra ; вентральная сегментарная область ; многие другие |
| Целевые ткани | По всей системе |
| Рецепторы | D 1 , D 2 , D 3 , D 4 , D 5 , Taar1 [ 3 ] |
| Агонисты | Direct: апоморфин , бромокриптин Косвенный : кокаин , амфетамин , метилфенидат |
| Антагонисты | Нейролептики , метаклопрамид , домперидон |
| Предшественник | Фенилаланин , тирозин и L-допа |
| Биосинтез | Dopa decarbobobroboboboxylase |
| Метаболизм | Мао , Комт [ 3 ] |
| Идентификаторы | |
| Номер CAS | |
| PubChem CID | |
| Iuphar/bps | |
| Наркоман | |
| Chemspider | |
| НЕКОТОРЫЙ | |
| Кегг | |
| Comptox Dashboard ( EPA ) | |
| Echa Infocard | 100.000.101 |
| Химические и физические данные | |
| Формула | C 8 H 11 N O 2 |
| Молярная масса | 153.181 g·mol −1 |
| 3D model ( JSmol ) | |
Дофамин ( DA , сокращение 3,4 D Ihydr O Xy Penhetimine , - ) представляет собой нейромодулирующую молекулу которая играет несколько важных ролей в клетках. Это органическое химическое вещество семейств катехоламинов и фенотиламинов . Дофамин составляет около 80% содержания катехоламинов в мозге. Это амин, синтезированный путем удаления карбоксильной группы из молекулы его химического вещества , L-допа , которая синтезируется в мозге и почках. Дофамин также синтезируется у растений и большинства животных. В мозге дофамин функционирует как нейротрансмиттер - химическое вещество, высвобождаемое нейронами (нервными клетками) для отправки сигналов в другие нервные клетки. Нейротрансмиттеры синтезируются в определенных областях мозга, но системно влияют на многие области. Мозг включает в себя несколько различных путей дофамина , один из которых играет важную роль в мотивационном компоненте поведения, мотивированного вознаграждением . Ожидание большинства типов вознаграждений повышает уровень дофамина в мозге, [ 4 ] и многие наркотики обратное увеличивают высвобождение дофамина или блокируют его захват в нейроны после высвобождения. [ 5 ] Другие пути дофамина мозга участвуют в моторном контроле и в контроле высвобождения различных гормонов. Эти пути и клеточные группы образуют дофаминовую систему, которая является нейромодулирующей . [ 5 ]
В популярной культуре и средствах массовой информации дофамин часто изображается как основное химическое вещество удовольствия, но нынешнее мнение в фармакологии заключается в том, что дофамин вместо этого придает мотивационную значимость ; [ 6 ] [ 7 ] [ 8 ] Другими словами, дофамин сигнализирует о воспринимаемой мотивационной известности (то есть желательности или отвращении) результата, который, в свою очередь, способствует поведению организма к или вне достижения этого результата. [ 8 ] [ 9 ]
За пределами центральной нервной системы дофамин функционирует в первую очередь как местный паракрин . В кровеносных сосудах он ингибирует высвобождение норэпинефрина и действует как вазодилататор ; В почках это увеличивает экскрецию натрия и выход мочи; В поджелудочной железе это уменьшает производство инсулина; В пищеварительной системе он снижает подвижность желудочно -кишечного тракта и защищает слизистую оболочку кишечника ; и в иммунной системе он снижает активность лимфоцитов . За исключением кровеносных сосудов, дофамин в каждой из этих периферических систем синтезируется локально и оказывает свои эффекты вблизи клеток, которые его высвобождают.
Несколько важных заболеваний нервной системы связаны с дисфункциями дофаминовой системы и некоторыми из ключевых лекарств, используемых для их работы, путем изменения эффектов дофамина. Болезнь Паркинсона , дегенеративное состояние, вызывающее тремор и моторные нарушения, вызвана потерей нейронов, секретирующих дофамин, в области среднего мозга, называемой чертикой субстанции . Его метаболический предшественник L-DOPA может быть изготовлен; Levodopa , чистая форма L-Dopa, является наиболее широко используемым лечением для Паркинсона. Существуют доказательства того, что шизофрения включает измененные уровни дофаминовой активности, а большинство антипсихотических препаратов, используемых для лечения этого, являются антагонистами дофамина , которые снижают активность дофамина. [ 10 ] Аналогичные препараты-антагонисты дофамина также являются одними из наиболее эффективных анти-наезе . Синдром беспокойных ног и дефицит внимания дефицита гиперактивности (СДВГ) связаны со снижением активности дофамина. [ 11 ] Дофаминергические стимуляторы могут быть привыкающими к высоким дозам, но некоторые используются в более низких дозах для лечения СДВГ. Сам дофамин доступен в качестве изготовленного лекарства от внутривенной инъекции . Это полезно при лечении тяжелой сердечной недостаточности или кардиогенного шока . [ 12 ] У новорожденных детей это может использоваться для гипотонии и септического шока . [ 13 ]
Структура
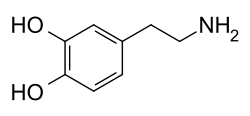
[ редактировать ]Молекула дофамина состоит из катехольной структуры ( бензоловое кольцо с двумя гидроксильных боковыми группами) с одной группой амина, прикрепленной через этильную цепь. [ 14 ] Таким образом, дофамин является самым простым возможным катехоламином , семейством, которое также включает нейротрансмиттеров норэпинефрина и адреналина . [ 15 ] Присутствие бензольного кольца с этим привязанностью амина делает его замещенным фенотиламином , семейством, которое включает в себя многочисленные психоактивные препараты . [ 16 ]
Как и большинство аминов, дофамин является органическим основанием . [ 17 ] В качестве основания он обычно протонируется в кислотных средах (в кислотно-основной реакции ). [ 17 ] Протонированная форма очень растворима в воде и относительно стабильной, но может стать окисленной, если она подвергается воздействию кислорода или других окислителей . [ 17 ] В основных средах дофамин не протонирован. [ 17 ] В этой свободной базовой форме он менее растворимый в воде, а также более реактивный. [ 17 ] Из-за повышенной стабильности и водопрощаемости протонированной формы дофамин поставляется для химического или фармацевтического использования в качестве дофамина гидрохлорида , то есть гидрохлоридной соли , которая создается, когда дофамин сочетается с гидрохлорной кислотой . [ 17 ] В сухой форме дофаминовый гидрохлорид представляет собой тонкий порошок, который из белого до желтого цвета. [ 18 ]


Биохимия
[ редактировать ]Биосинтетические пути для катехоламинов и просвечивания аминов в человеческом мозге [ 19 ] [ 20 ] [ 21 ]
|
Синтез
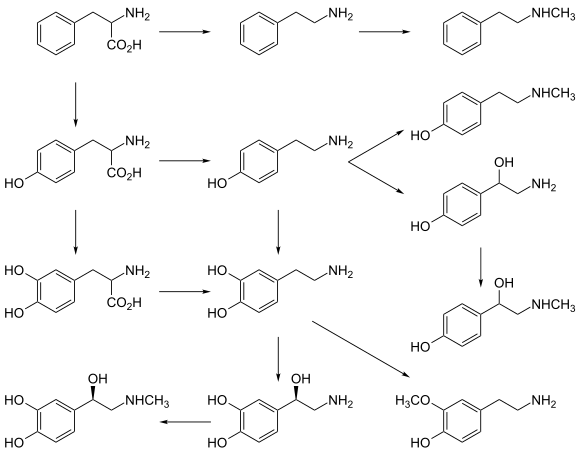
[ редактировать ]Дофамин синтезируется в ограниченном наборе типов клеток, в основном нейронов и клеток мозге надпочечников в . [ 22 ] Основными и незначительными метаболическими путями соответственно являются:
- Первичный: L -фенилаланин → L -тирозин → L -допа → дофамин [ 19 ] [ 20 ]
- Минор: L -фенилаланин → L -тирозин → P -тирамин → дофамин [ 19 ] [ 20 ] [ 21 ]
- Минор: L -фенилаланин → M -тирозин → M -тирамин → дофамин [ 21 ] [ 23 ] [ 24 ]
Прямой предшественник дофамина, L -допа , может быть косвенно синтезирован от незаменимых аминокислотных фенилаланина или непосредственно из несущественного аминокислотного тирозина . [ 25 ] Эти аминокислоты встречаются почти в каждом белке и поэтому легко доступны в пище, причем тирозин является наиболее распространенным. Хотя дофамин также обнаруживается во многих типах пищи, он не способен пересечь барьер крови -мозга , который окружает и защищает мозг. [ 26 ] Поэтому он должен быть синтезирован внутри мозга, чтобы выполнить свою нейрональную активность . [ 26 ]
L -фенилаланин превращается в L -тирозин ферментом фенилаланиновой гидроксилазы с молекулярным кислородом (O 2 ) и тетрагидробиоптерином в качестве кофакторов . L -тирозин превращается в L -допа ферментной тирозин -гидроксилазой с тетрагидробиоптерином, O 2 и железом (Fe 2+ ) как кофакторы. [ 25 ] L -допа превращается в дофамин ферментом ароматической л -аминокислот декарбоксилазы (также известной как допа -декарбоксилаза), с пиридоксаль -фосфатом в качестве кофактора. [ 25 ]
Сам дофамин используется в качестве предшественника в синтезе нейротрансмиттеров норэпинефрина и адреналина. [ 25 ] Дофамин превращается в норэпинефрин ферментом дофамин β -гидроксилазы , с O 2 и L -аскорбиновой кислотой в качестве кофакторов. [ 25 ] Норепинефрин превращается в адреналин ферментом фенилэтаноламин N -метилтрансферазы с S -аденозил -л -метионином в качестве кофактора. [ 25 ]
Некоторые из кофакторов также требуют собственного синтеза. [ 25 ] Дефицит в любой необходимой аминокислоте или кофакторе может ухудшить синтез дофамина, норэпинефрина и адреналина. [ 25 ]
Деградация
[ редактировать ]Дофамин разбивается на неактивные метаболиты с помощью набора ферментов - моноаминоксидазы (MAO), катехол -о -метилтрансфераза (COMT) и альдегиддегидрогеназы (ALDH), действуя в последовательности. [ 27 ] Обе изоформы моноаминоксидазы, MAO-A и MAO-B , эффективно метаболизируют дофамин. [ 25 ] Существуют различные пути разрушения, но основным конечным продуктом является гомованиллиновая кислота (HVA), которая не имеет известной биологической активности. [ 27 ] Из кровотока гомованиллиновая кислота фильтруется почками, а затем выделяется в моче. [ 27 ] Два первичных метаболических маршрута, которые преобразуют дофамин в HVA: [ 28 ]
- Дофамин → допал → Dopac → HVA - катализируется MAO, ALDH и COMT соответственно
- Дофамин → 3-метокситирамин → HVA-катализируется COMT и MAO+ALDH соответственно
В клинических исследованиях шизофрении измерения гомованиллической кислоты в плазме использовались для оценки уровней активности дофамина в мозге. Однако сложность в этом подходе заключается в разделении высокого уровня гомованиллической кислоты в плазме, вносящем метаболизм норэпинефрина. [ 29 ] [ 30 ]
Хотя дофамин обычно разрушается оксидоредуктазой , он также подвержен окислению прямой реакцией с кислородом, давая хиноны плюс различные свободные радикалы в качестве продуктов. [ 31 ] Скорость окисления может быть увеличена за счет присутствия железа железа или других факторов. Хиноны и свободные радикалы, продуцируемые путем автооксидирования дофамина, могут отравить ядовитые клетки , и есть доказательства того, что этот механизм может способствовать потерь клеток, возникающих при болезни Паркинсона и других состояниях. [ 32 ]
Функции
[ редактировать ]Клеточные эффекты
[ редактировать ]| Семья | Рецептор | Ген | Тип | Механизм |
|---|---|---|---|---|
| D 1 -like | D 1 | DRD1 | G s -куплент. | Увеличить внутриклеточные уровни лагеря Активацией аденилатциклазы . |
| D 5 | DRD5 | |||
| D 2 -like | D 2 | DRD2 | G I -Coupled. | Уменьшить внутриклеточные уровни лагеря ингибируя аденилатциклазу . |
| D 3 | DRD3 | |||
| D 4 | DRD4 | |||
| Простой | Таар1 | Таар1 | G s -куплент. G Q -Куплент. |
Увеличить внутриклеточные уровни лагеря и внутриклеточная концентрация кальция. |
Дофамин оказывает свое воздействие, связывая и активируя рецепторы клеточной поверхности . [ 22 ] У людей дофамин обладает высокой аффинностью связывания в рецепторах дофамина и рецепторе 1-амина человека (HTAAR1). [ 3 ] [ 33 ] У млекопитающих пять подтипов дофаминовых рецепторов было идентифицировано , помеченных от D 1 до D 5 . [ 22 ] Все они функционируют как метаботропные , G-белковые рецепторы , что означает, что они оказывают свои эффекты через сложную систему второго мессенджера . [ 34 ] Эти рецепторы могут быть разделены на две семьи, известные как D 1 -похожи и D 2 -похожи . [ 22 ] Для рецепторов, расположенных на нейронах в нервной системе, окончательным эффектом D 1 -подобной активации (D 1 и D 5 ) может быть возбуждение (посредством открытия натриевых каналов ) или ингибирование (посредством открытия каналов калия ); Конечный эффект D 2 -подобной активации (D 2 , D 3 и D 4 ) обычно ингибирует нейрона -мишени. [ 34 ] Следовательно, неверно описывать сам дофамин как возбуждающий или ингибирующий: его влияние на нейрона -мишени зависит от того, какие типы рецепторов присутствуют на мембране этого нейрона и от внутренних реакций этого нейрона на второй лагерь мессенджера . [ 34 ] D 1 Рецепторы являются наиболее многочисленными дофаминовыми рецепторами в нервной системе человека; D 2 Рецепторы следующие; D 3 , D 4 и D 5 Рецепторы присутствуют на значительно более низких уровнях. [ 34 ]
Хранение, выпуск и обратный захват
[ редактировать ]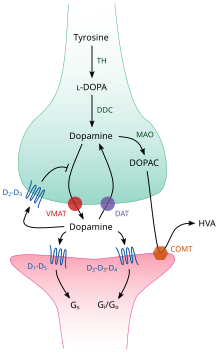
TH: тирозин -гидроксилаза
Dopa: L-Dopa
Это: дофаминовый транспортер
DDC: Dopa Decarbobobrobobobrobobroboboboxylase
VMAT: везикулярный моноаминовый транспортер 2
Мао: моноаминоксидаза
COMT: Catechol-O-метилтрансфераза
Что: гомованиллическая кислота
Внутри мозга дофамин функционирует как нейротрансмиттер и нейромодулятор и контролируется набором механизмов, общих для всех моноротрансмиттеров . [ 22 ] После синтеза дофамин транспортируется из цитозоля в синаптические везикулы растворителем растворенного носителя - везикулярный транспортер моноамин , VMAT2 . [ 35 ] Дофамин хранится в этих везикулах до тех пор, пока он не будет выброшен в синаптическую расщелину . В большинстве случаев высвобождение дофамина происходит посредством процесса, называемого экзоцитозом , который вызван потенциалами действия , но также может быть вызвана активностью внутриклеточного рецептора, ассоциированного с амином , TAAR1 . [ 33 ] TAAR1 является высокоаффинным рецептором для дофамина, следов аминов и некоторых замещенных амфетаминов , которые расположены вдоль мембран во внутриклеточной среде пресинаптической клетки; [ 33 ] Активация рецептора может регулировать передачу сигналов дофамина, индуцируя ингибирование и отток дофамина и отток , а также путем ингибирования стрельбы нейронов через разнообразный набор механизмов. [ 33 ] [ 36 ]
Оказавшись в синапсе, дофамин связывается и активирует дофаминовые рецепторы. [ 37 ] Это могут быть постсинаптические дофаминовые рецепторы, которые расположены на дендритах (постсинаптический нейрон) или пресинаптических авторецепторов (например, D 2 Sh и пресинаптический D 3 -рецепторы), которые расположены на мембране терминала аксона (пресинаптический нейрон), которые расположены на мембране терминала аксона (пресинаптический нейрон), которые расположены на мембране терминала аксона (пресинаптический нейрон), которые расположены на мембране терминала аксона (пресинаптический нейрон), которые расположены на мембране терминала аксона (пресинаптический нейрон), которые расположены на мембране терминала аксонов (пресинаптический нейрон), которые расположены на мембране терминала аксона (пресинаптический нейрон)) Полем [ 22 ] [ 37 ] После того, как постсинаптический нейрон вызывает потенциал действия, молекулы дофамина быстро становятся несвязанными от их рецепторов. Затем они поглощаются обратно в пресинаптическую клетку, посредством обратного захвата, опосредованного либо транспортером дофамина , либо транспортером моноаминов плазматической мембраны . [ 38 ] Вернувшись в цитозоль, дофамин может быть разбит либо моноаминоксидазой , либо перепакован в пузырьки VMAT2, что делает его доступным для будущего выпуска. [ 35 ]
В мозге уровень внеклеточного дофамина модулируется двумя механизмами: фазическая и тоническая передача . [ 39 ] Фазическое высвобождение дофамина, как и большинство высвобождения нейротрансмиттера в нервной системе, управляется непосредственно потенциалами действия в дофаминосодержащих клетках. [ 39 ] Тонизированное дофаминовое передача происходит, когда небольшие количества дофамина высвобождаются без предшествующих потенциалов пресинаптического действия. [ 39 ] Тоническая передача регулируется различными факторами, включая активность других нейронов и обратного захвата нейротрансмиттера. [ 39 ]
Центральная нервная система
[ редактировать ]
Внутри мозга дофамин играет важную роль в исполнительных функциях , моторном контроле , мотивации , возбуждении , подкреплении и вознаграждении , а также на функциях более низкого уровня, включая лактацию , сексуальное удовлетворение и тошноту . Группы дофаминергических клеток и пути составляют дофаминовую систему, которая является нейромодулирующей .
Дофаминергические нейроны (дофамин-продуцирующие нервные клетки) сравнительно мало-в общей сложности около 400 000 в мозге человека [ 40 ] - и их клеточные тела ограничены группами несколькими относительно небольшими областями мозга. [ 41 ] Однако их аксоны проецируются во многих других областях мозга, и они оказывают сильное влияние на свои цели. [ 41 ] Эти группы дофаминергических клеток были впервые нанесены на карту в 1964 году Анникой Далстрём и Кьеллом Фусэ, которые назначили им этикетки, начиная с буквы «A» (для «аминергического»). [ 42 ] В своей схеме участки A1 до A7 содержат нейротрансмиттер Norepinephrin, тогда как A8 до A14 содержат дофамин. Дофаминергические районы, которые они идентифицировали, - это черная субстанция (группы 8 и 9); вентральная область сегмента (группа 10); задний гипоталамус (группа 11); дугообразное ядро (группа 12); Zona Incerta (группа 13) и перивентрикулярное ядро (группа 14). [ 42 ]
Субъективная привязка - это небольшая область среднего мозга, которая образует компонент базальных ганглиев . Это имеет две части - область ввода, называемую ретикулатами PARS , и площадь вывода, называемая PARS Compacta . Дофаминергические нейроны обнаруживаются в основном в PARS Compacta (клеточная группа A8) и поблизости (группа A9). [ 41 ] У людей проекция дофаминергических нейронов от субстанции nigra pars compacta до дорсального стриатума, называемой нигростриатальным путем , играет значительную роль в контроле моторных функций и в изучении новых моторных навыков . [ 43 ] Эти нейроны особенно уязвимы для повреждения, и когда большое количество из них умирает, результатом является синдром паркинсона . [ 44 ]
Вентральная сегментарная область (VTA) является еще одной областью среднего мозга. Наиболее выдающаяся группа дофаминергических нейронов VTA проецирует в префронтальную кору через мезокортикальный путь и еще одну меньшую группу проектов в ядро, прилежащем через мезолимбический путь . Вместе эти два пути коллективно называются мезокортиколимбической проекцией . [ 41 ] [ 43 ] VTA также посылает дофаминергические проекции в миндалину , поясную извилину , гиппокамп и обонятельную лампу . [ 41 ] [ 43 ] Мезокортиколимбические нейроны играют центральную роль в награде и других аспектах мотивации. [ 43 ] Накопление литературы показывает, что дофамин также играет решающую роль в аверсивном обучении через его влияние на ряд областей мозга. [ 45 ] [ 46 ] [ 47 ]
Задний гипоталамус имеет дофаминовые нейроны, которые проецируются в спинном мозге, но их функция не очень хорошо установлена. [ 48 ] Есть некоторые доказательства того, что патология в этой области играет роль в синдроме беспокойных ног, состояние, при котором люди испытывают трудности со сном из -за подавляющего принуждения к постоянному перемещению частей тела, особенно ног. [ 48 ]
Ядро дугообразных и перивентрикулярное ядро гипоталамуса имеют дофаминовые нейроны, которые образуют важную проекцию - тубероинфункциональный путь , который идет в гипофиз , где он влияет на секрецию гормона пролактина . [ 49 ] Дофамин является основным нейроэндокринным ингибитором секреции пролактина из передней гипофизы. [ 49 ] Дофамин, продуцируемый нейронами в дугообразном ядре, секретируется в гипофизарную портальную систему среднего возвышения , которая поставляет гипофиз . [ 49 ] Пролактиновые клетки , которые продуцируют пролактин, в отсутствие дофамина непрерывно секретируют пролактин; Дофамин ингибирует эту секрецию. [ 49 ]
Zona Incerta, сгруппированная между дугообразными и перивентрикулярными ядрами, проецирует несколько областей гипоталамуса, и участвует в контроле гонадотропин-релезингового гормона , который необходим для активации развития мужчин и женщин репродуктивных систем , после полового созревания. [ 49 ]
Дополнительная группа нейронов, секретирующих дофамин, обнаружена в сетчатке глаза. [ 50 ] Эти нейроны являются амакринными клетками , что означает, что у них нет аксонов. [ 50 ] Они высвобождают дофамин во внеклеточную среду и специально активны в дневное время, молча о том, как ночью молчит. [ 50 ] Этот дофамин сетчатки действует для усиления активности конусных клеток в сетчатке при подавлении клеток стержня - результатом является повышение чувствительности к цвету и контрастности в условиях яркого освещения, за счет снижения чувствительности, когда свет туст. [ 50 ]
Базальные ганглии
[ редактировать ]
Крупнейшими и наиболее важными источниками дофамина в мозге позвоночных являются черная субстанция и вентральная сегментарная область. [ 41 ] Обе структуры являются компонентами среднего мозга, тесно связаны друг с другом и во многих отношениях функционально сходны. [ 41 ] Крупнейшим компонентом базальных ганглиев является стриатум. [ 51 ] Применение субстанции посылает дофаминергическую проекцию в дорсальный стриатум , в то время как вентральная сегментарная область посылает аналогичный тип дофаминергической проекции в вентральный стриатум . [ 41 ]
Прогресс в понимании функций базальных ганглиев был медленным. [ 51 ] Наиболее популярные гипотезы, широко заявленные, предлагают, чтобы базальные ганглии играли центральную роль в выборе действий . [ 52 ] Теория отбора действий в ее простейшей форме предполагает, что, когда человек или животное находятся в ситуации, когда возможно несколько поведений, активность в базальных ганглиях определяет, кто из них выполняется, путем освобождения этого ответа из ингибирования, продолжая ингибировать другие двигательные системы что если активировать будет генерировать конкурирующее поведение. [ 53 ] Таким образом, базальные ганглии в этой концепции ответственны за инициирование поведения, но не за определение деталей того, как они выполняются. Другими словами, они по сути формируют систему принятия решений. [ 53 ]
Базальные ганглии можно разделить на несколько секторов, и каждый участвует в контроле определенных типов действий. [ 54 ] Вентральный сектор базальных ганглиев (содержащий вентральный стриатум и вентральный сегментар) работает на самом высоком уровне иерархии, выбирая действия на уровне всего организатора. [ 53 ] Спинные сектора (содержащие дорсальный стриатум и черновая субстанция) работают на более низких уровнях, выбирая специфические мышцы и движения, которые используются для реализации данной схемы поведения. [ 54 ]
Дофамин вносит вклад в процесс выбора действий по крайней мере двумя важными способами. Во -первых, он устанавливает «порог» для инициирования действий. [ 52 ] Чем выше уровень дофаминовой активности, тем ниже импульс, необходимый для вызывания данного поведения. [ 52 ] Как следствие, высокий уровень дофамина приводит к высоким уровням двигательной активности и импульсивного поведения ; Низкие уровни дофамина приводят к уплотнению и замедляли реакции. [ 52 ] Болезнь Паркинсона, при которой уровни дофамина в цепи субстанции применения значительно снижаются, характеризуется жесткостью и затруднением, инициирующим движение - однако, когда люди с болезнью сталкиваются с сильными стимулами, такими как серьезная угроза, их реакции могут быть такими же энергичными, как и жизне те из здорового человека. [ 55 ] В противоположном направлении лекарства, которые увеличивают высвобождение дофамина, такие как кокаин или амфетамин, могут вызывать повышенные уровни активности, в том числе в крайнем психомоторном агитации и стереотипных движениях . [ 56 ]
Вторым важным эффектом дофамина является «обучающий» сигнал. [ 52 ] Когда действие сопровождается увеличением активности дофамина, схема базальных ганглиев изменяется таким образом, что облегчает тот же ответ, чтобы вызвать, когда в будущем возникнут подобные ситуации. [ 52 ] Это форма оперантной кондиционирования , в которой дофамин играет роль сигнала вознаграждения. [ 53 ]
Награда
[ редактировать ]
На языке, используемом для обсуждения системы вознаграждения, вознаграждение является привлекательным и мотивационным свойством стимула, которое вызывает аппетитное поведение (также известное как поведение подхода) и консультанное поведение . [ 57 ] Получительный стимул - это тот, который может побудить организм приближаться к нему и выбрать его. [ 57 ] Удовольствие , обучение (например, классическая и оперантная кондиционирование ) и поведение подхода являются тремя основными функциями вознаграждения. [ 57 ] Как аспект вознаграждения, удовольствие дает определение вознаграждения; [ 57 ] Однако, хотя все приятные стимулы полезны, не все полезные стимулы приятны (например, внешние награды, такие как деньги). [ 57 ] [ 58 ] Мотивационный или желательный аспект полезных стимулов отражается в поведении подхода, которое они вызывают, тогда как удовольствие от внутренних вознаграждений является результатом употребления их после их приобретения. [ 57 ] Нейропсихологическая модель, которая различает эти два компонента необычайно полезного стимула, является моделью стимулирующей значимости , где «желание» или желание (реже, «поиск» [ 59 ] ) соответствует аппетитному или подходному поведению, в то время как «симпатия» или удовольствие соответствует контумируемому поведению. [ 57 ] [ 6 ] [ 60 ] У наркоманов человека «желание» растет с «симпатии», поскольку желание использовать привыкание препарата увеличивается, в то время как удовольствие, полученное от потребления его, уменьшается из -за толерантности к наркотикам . [ 6 ]
Внутри мозга дофамин функционирует частично как глобальный сигнал вознаграждения. Первоначальный ответ дофамина на полезный стимул кодирует информацию о значимости , ценности и контексте вознаграждения. [ 57 ] В контексте обучения, связанного с вознаграждением, дофамин также функционирует как сигнал ошибки прогнозирования вознаграждения , то есть степень, в которой значение вознаграждения неожиданно. [ 57 ] Согласно этой гипотезе, предложенной Монтегью, Даяном и Седжновски, [ 61 ] Награды, которые ожидаются, не проводят второй фазовый дофаминовый ответ в определенных дофаминергических клетках, но награды, которые являются неожиданными или больше, чем ожидалось, вызывают краткосрочное увеличение синаптического дофамина, тогда как упущение ожидаемого вознаграждения на самом деле вызывает высвобождение дофамина. упасть ниже его фонового уровня. [ 57 ] Гипотеза «ошибка прогнозирования» вызвала особый интерес от вычислительных нейробиологов, потому что влиятельный метод вычислительного обучения, известный как обучение временной разнице, обеспечивает сильное использование сигнала, который кодирует ошибку прогнозирования. [ 57 ] Это слияние теории и данных привело к плодотворному взаимодействию между нейробиологами и компьютерными учеными, заинтересованными в машинном обучении . [ 57 ]
Данные из записей микроэлектродов из мозга животных показывают, что дофаминовые нейроны в вентральной сегментарной области (VTA) и черновой субстанции сильно активируются широким разнообразием полезных событий. [ 57 ] Эти чувствительные к вознаграждению дофаминовые нейроны в VTA и субстанции имеют решающее значение для познания, связанных с вознаграждением и служат центральным компонентом системы вознаграждения. [ 6 ] [ 62 ] [ 63 ] Функция дофамина варьируется в каждой аксональной проекции от VTA и субстанции NIGRA; [ 6 ] Например, проекция оболочки ucleus accumbens accumbens присвоена стимулирующей значимостью («хочу») для вознаграждения стимулов и связанных с ними сигналов , проекции префронтальной коры VTA - префронтальной коры обновляет ценность различных целей в соответствии с их стимулирующей значимостью, VTA -Amygdala и Проекции VTA-Hippocampus опосредуют консолидацию воспоминаний, связанных с вознаграждением, и как ядра VTA-ядра, прилегающее ядро , так и черновые пути субстанции. [ 6 ] [ 64 ] Некоторая активность в дофаминергических проекциях VTA, по -видимому, также связана с прогнозом вознаграждения. [ 6 ] [ 64 ]
Удовольствие
[ редактировать ]В то время как дофамин играет центральную роль в выборе «желания», связанного с аппетитным или подходящим поведенческим реакцией к полезным стимулам, подробные исследования показали, что дофамин не может быть просто приравнивается к гедонистическому «симпатию» или удовольствию, что отражено в консультантом поведенческом ответе. [ 58 ] Нейротрансмиссия дофамина участвует во всех, но не во всех аспектах связанных с удовольствием познания, поскольку центры удовольствия были идентифицированы как в системе дофамина (то есть, ядро прикупаемость), так и за пределами дофаминовой системы (т.е. вентральное паллидум и парабочевое ядро ). [ 58 ] [ 60 ] [ 65 ] Например, прямая электрическая стимуляция дофаминовых путей с использованием электродов, имплантированных в мозг, воспринимается как приятная, и многие виды животных готовы работать, чтобы получить его. [ 66 ] Антипсихотические препараты снижают уровень дофамина и, как правило, вызывают анхедонию , уменьшенную способность испытывать удовольствие. [ 67 ] Многие виды приятных переживаний, такие как половые акты, еда и игра в видеоигры, - воспитание выпуска дофамина. [ 68 ] Все привыкающие препараты прямо или косвенно влияют на нейротрансмиссию дофамина в прилежащем ядре; [ 6 ] [ 66 ] Эти препараты увеличивают «желание», что приводит к компульсивному употреблению лекарств, когда он неоднократно принимал в высоких дозах, предположительно благодаря сенсибилизации стимулирования . [ 60 ] Препараты, которые увеличивают концентрации синаптического дофамина, включают психостимуляторы , такие как метамфетамин и кокаин. Они вызывают увеличение поведения «желания», но не очень изменяют выражения удовольствия или уровня изменений насыщения. [ 60 ] [ 66 ] Тем не менее, опиатные препараты, такие как героин и морфин, увеличивают выражения «симпатии» и «желания» поведения. [ 60 ] Более того, животные, у которых вентральная сегментарная система дофаминовой системы стала неактивной, не ищут пищи, и будут голодать до смерти, если оставят себе, но если пища помещается в их рты, они будут потреблять ее и показывать выражения, указывающие на удовольствие. [ 69 ]
Клиническое исследование с января 2019 года, в котором оценивалось влияние предшественника дофамина ( леводопа ), антагониста допамина ( рисперидон ) и плацебо на реакции вознаграждения на музыку - включая степень удовольствия, испытываемого во время музыкальных озноб , измеряемые изменениями в электродермальной активности. а также субъективные оценки - обнаружили, что манипуляция с нейротрансмиссией дофамина двунаправленно регулирует познание удовольствия (в частности, гедоническое воздействие музыки ) у людей. [ 70 ] [ 71 ] Это исследование продемонстрировало, что повышенная нейротрансмиссия дофамина действует как нерабочее состояние для приятных гедонических реакций на музыку у людей. [ 70 ] [ 71 ]
Исследование, опубликованное в природе в 1998 году, обнаружило доказательства того, что игра в видеоигры выпускает дофамин в человеческом стриатуме. Этот дофамин связан с обучением, подкреплением поведения, вниманием и сенсомоторной интеграцией. [ 72 ] Исследователи использовали сканирование позитронной эмиссионной томографии и 11 C-метеобороновый растист для отслеживания уровней дофамина в мозге во время целенаправленных моторных задач и обнаружил, что высвобождение дофамина положительно коррелировало с выполнением задач и был наибольшим в вентральном стриатуме . Это было первое исследование, которое продемонстрировало поведенческие условия, при которых дофамин выпускается у людей. Он подчеркивает способность позитронной эмиссионной томографии обнаруживать потоки нейротрансмиттеров во время изменений в поведении. Согласно исследованию, потенциально проблематичное использование видеоигр связано с такими чертами личности, как низкая самооценка и низкая самоэффективность, тревога, агрессия и клинические симптомы депрессии и тревожные расстройства. [ 73 ] Кроме того, причины, по которым люди играют в видеоигры, различаются и могут включать в себя преодоление , социализацию и личное удовлетворение. DSM -5 определяет расстройство интернет-игр как психическое расстройство, тесно связанное с расстройством азартных игр. Это было поддержано некоторыми исследователями, но также вызвало противоречие.
За пределами центральной нервной системы
[ редактировать ]Дофамин не пересекает барьер крови -мрака, поэтому его синтез и функции в периферических областях в значительной степени не зависят от его синтеза и функций в мозге. [ 26 ] Значительное количество дофамина циркулирует в кровотоке, но его функции там не совсем ясны. [ 27 ] Дофамин обнаруживается в плазме крови на уровнях, сопоставимых с уровнями адреналина, но у людей более 95% дофамина в плазме расположены в виде дофамина сульфата , конъюгат, продуцируемого ферментом сульфортрансферазы 1A3/1A4, действующего на свободный дофамин. [ 27 ] Большая часть этого дофамина сульфата производится в брыжеечных органах. [ 27 ] Считается, что производство дофамина сульфата является механизмом детоксифицирующего дофамина, который принимается в качестве пищи или производится в результате пищеварительного процесса-выросшие в плазме обычно возрастают более чем в пятьдесят раз после еды. [ 27 ] Сульфат дофамина не имеет известных биологических функций и выводится в моче. [ 27 ]
Относительно небольшое количество неконъюгированного дофамина в кровотоке может быть получена симпатической нервной системой , пищеварительной системой или, возможно, другими органами. [ 27 ] Он может действовать на дофаминовых рецепторах в периферических тканях или метаболизируется, или быть превращенным в норэпинефрин ферментом дофаминовой бета -гидроксилазы , которая высвобождается в кровоточаще медуллой надпочечников. [ 27 ] Некоторые дофаминовые рецепторы расположены в стенках артерий, где они действуют как вазодилататор и ингибитор высвобождения норэпинефрина из терминалов постганглионного симпатических нервов (дофамин может ингибировать норпинефрино. как сама норэпинефрин). [ 74 ] Эти ответы могут быть активированы дофамином, выделяемым из тела сонной артерии в условиях низкого кислорода, но не известны ли артериальные дофаминовые рецепторы. [ 74 ]
Помимо его роли в модулировании кровотока, существует несколько периферических систем, в которых дофамин циркулирует в пределах ограниченной области и выполняет экзокрину или паракринную функцию. [ 27 ] Периферические системы, в которых дофамин играет важную роль, включают иммунную систему , почки и поджелудочную железу .
Иммунная система
[ редактировать ]В иммунной системе дофамин действует на рецепторы, присутствующие на иммунных клетках, особенно лимфоцитах . [ 75 ] Дофамин также может влиять на иммунные клетки в системе селезенки , костного мозга и кровообращения . [ 76 ] Кроме того, дофамин может быть синтезирован и выделен самими иммунными клетками. [ 75 ] Основным эффектом дофамина на лимфоциты является снижение уровня их активации. Функциональная значимость этой системы неясно, но она дает возможный путь для взаимодействия между нервной системой и иммунной системой и может иметь отношение к некоторым аутоиммунным расстройствам. [ 76 ]
Почки
[ редактировать ]Почечная дофаминергическая система расположена в клетках нефрона в почке, где присутствуют все подтипы дофаминовых рецепторов. [ 77 ] Дофамин также синтезируется там, с помощью клеток для канальцев и разгружается в трубчатую жидкость . Его действия включают в себя увеличение кровоснабжения в почках, увеличение скорости клубочковой фильтрации и увеличение экскреции натрия в моче. Следовательно, дефекты в почечной функции дофамина могут привести к снижению экскреции натрия и, следовательно, привести к развитию высокого кровяного давления . Существуют убедительные доказательства того, что неисправности в производстве дофамина или в рецепторах могут привести к ряду патологий, включая окислительный стресс , отек и генетическую или важную гипертонию. Окислительный стресс сам по себе может вызвать гипертонию. [ 78 ] Дефекты в системе также могут быть вызваны генетическими факторами или высоким кровяным давлением. [ 79 ]
Поджелудочная железа
[ редактировать ]В поджелудочной железе роль дофамина несколько сложна. Поджелудочная железа состоит из двух частей, экзокринных и эндокринных компонентов. Экзокринная часть синтезирует и секретирует пищеварительные ферменты и другие вещества, включая дофамин, в тонкую кишку. [ 80 ] Функция этого секретируемого дофамина после того, как он входит в тонкую кишку, не установлена четко - возможности включают защиту слизистой оболочки кишечника от повреждения и снижение подвижности желудочно -кишечного тракта (скорость, с которой содержимое движется через пищеварительную систему). [ 80 ]
Островки поджелудочной железы составляют эндокринную часть поджелудочной железы и синтезируют и секретируют гормоны, включая инсулин в кровоток. [ 80 ] Существуют доказательства того, что бета -клетки на островках, которые синтезируют инсулин, содержат дофаминовые рецепторы, и что дофамин действует для уменьшения количества инсулина, который они высвобождают. [ 80 ] Источник их дофаминового ввода не установлен четко - он может исходить из дофамина, который циркулирует в кровотоке и вытекает из симпатической нервной системы, или он может быть синтезирован локально другими типами клеток поджелудочной железы. [ 80 ]
Медицинское использование
[ редактировать ]
Дофамин как промышленное лекарство продается под торговыми названиями, вставными, допастатами и ревминами. Это в списке основных лекарств Всемирной организации здравоохранения . [ 81 ] Он чаще всего используется в качестве стимулирующего препарата при лечении тяжелого низкого кровяного давления , медленной частоты сердечных сокращений и остановки сердца . Это особенно важно при их лечении у новорожденных . [ 82 ] [ 13 ] Это дается внутривенно. Поскольку период полураспада дофамина в плазме очень короткий-примерно одна минута у взрослых, две минуты у новорожденных и до пяти минут у недоношенных детей-обычно дается в непрерывной внутривенной капельнице, а не в одной инъекции. [ 83 ]
Его эффекты, в зависимости от дозировки, включают в себя увеличение экскреции натрия по почках, увеличение мочи, увеличение частоты сердечных сокращений и увеличение артериального давления . [ 83 ] В низких дозах он действует через симпатическую нервную систему, чтобы увеличить силу сокращения сердечных мышц и частоту сердечных сокращений, тем самым увеличивая сердечный выброс и артериальное давление. [ 84 ] Более высокие дозы также вызывают вазоконстрикцию , которая еще больше повышает артериальное давление. [ 84 ] [ 85 ] Старая литература также описывает очень низкие дозы, которые, как считаются, улучшают функцию почек без других последствий, но недавние обзоры пришли к выводу, что дозы на таких низких уровнях не являются эффективными и иногда могут быть вредными. [ 86 ] В то время как некоторые эффекты являются результатом стимуляции дофаминовых рецепторов, заметные сердечно -сосудистые эффекты являются результатом дофамина, действующего в α 1 , β 1 и β 2 адренергических рецепторов β и β 2 . [ 87 ] [ 88 ]
Побочные эффекты дофамина включают негативное влияние на функцию почек и нерегулярное сердцебиение . [ 84 ] Было обнаружено, что доза LD 50 , или летальная доза, которая, как ожидается, окажется смертельной для 50% населения: 59 мг/кг (мышь; вводится внутривенно ); 95 мг/кг (мышь; вводятся внутрибрюшинно ); 163 мг/кг (крыса; вводятся внутрибрюшинно); 79 мг/кг (собака; вводится внутривенно). [ 89 ]
Болезнь, расстройства и фармакология
[ редактировать ]Система дофамина играет центральную роль в нескольких значительных медицинских состояниях, включая болезнь Паркинсона , дефицит внимания гиперактивность , синдром Туретта , шизофрении , биполярное расстройство и зависимость . Помимо самого дофамина, существует много других важных препаратов, которые действуют на дофаминовые системы в различных частях мозга или тела. Некоторые используются в медицинских или рекреационных целях, но нейрохимики также разработали различные исследовательские препараты, некоторые из которых связываются с высокой аффинностью с конкретными типами рецепторов дофамина и либо агонизируют , либо противодействуют их последствиям, и многие, которые влияют на другие аспекты физиологии дофамина В [ 90 ] включая ингибиторы транспортера дофамина , ингибиторы VMAT и ингибиторы фермента .
Старение мозга
[ редактировать ]В ряде исследований сообщалось о возрастном снижении синтеза дофамина и плотности дофаминовых рецепторов (то есть количество рецепторов) в мозге. [ 91 ] Было показано, что этот упадок происходит в стриатуме и экстрастриатских регионах. [ 92 ] Уменьшение рецепторов D 1 , D 2 и D 3 хорошо задокументировано. [ 93 ] [ 94 ] [ 95 ] Считается, что снижение дофамина с старением отвечает за многие неврологические симптомы, которые увеличиваются в частоте с возрастом, такие как снижение качания рук и повышенная жесткость . [ 96 ] Изменения в уровнях дофамина также могут вызвать возрастные изменения в когнитивной гибкости. [ 96 ]
Рассеянный склероз
[ редактировать ]Исследования показали, что дисбаланс дофамина влияет на усталость при рассеянном склерозе . [ 97 ] У пациентов с рассеянным склерозом дофамин ингибирует продукцию IL-17 и IFN-γ мононуклеарными клетками периферической крови. [ 98 ]
Болезнь Паркинсона
[ редактировать ]Болезнь Паркинсона-это возрастное расстройство, характеризующееся расстройствами движения , такими как жесткость тела, замедление движения и дрожь конечности, когда они не используются. [ 55 ] На продвинутых этапах он переходит к деменции и в конечном итоге смертью. [ 55 ] Основные симптомы вызваны потерей дофаминовых секретирующих клеток в черной субстанции. [ 99 ] Эти дофаминовые клетки особенно уязвимы для повреждений, и множество оскорблений, включая энцефалит (как изображено в книге и пробуждениях спорта к фильмам), повторные сотрясения и некоторые формы химического отравления, такие как MPTP , могут привести к существенной потере клеток , создавая синдром паркинсона , который похож на его основные особенности с болезнью Паркинсона. [ 100 ] Однако большинство случаев болезни Паркинсона являются идиопатическими , что означает, что причина гибели клеток не может быть идентифицирована. [ 100 ]
Наиболее широко используемым лечением для паркинсонизма является введение L-DOPA, предшественник метаболического дофамина. [ 26 ] L-DOPA is converted to dopamine in the brain and various parts of the body by the enzyme DOPA decarboxylase. [ 25 ] L-DOPA используется, а не сам дофамин, потому что, в отличие от дофамина, он способен пересечь барьер кровь-мран . [ 26 ] Он часто проводит совместное управление с ингибитором фермента периферического декарбоксилирования , такого как карбидопа или бенсеразид , чтобы уменьшить количество, преобразованное в дофамин в периферии, и тем самым увеличивать количество L-DOPA, которое попадает в мозг. [ 26 ] Когда L-DOPA регулярно вводится в течение длительного периода времени, различные неприятные побочные эффекты, такие как дискинезия , часто начинают появляться; Несмотря на это, это считается лучшим доступным долгосрочным вариантом лечения для большинства случаев болезни Паркинсона. [ 26 ]
Обработка L-DOPA не может восстановить дофаминовые клетки, которые были потеряны, но приводит к тому, что оставшиеся клетки продуцируют больше дофамина, тем самым компенсируя потерю по меньшей мере до некоторой степени. [ 26 ] На продвинутых этапах лечение начинает терпеть неудачу, потому что потери клеток настолько сильны, что оставшиеся не могут производить достаточное количество дофамина независимо от уровней L-DOPA. [ 26 ] Другие препараты, которые усиливают функцию дофамина, такие как бромокриптин и перголид , также иногда используются для лечения паркинсонизма, но в большинстве случаев L-DOPA, по-видимому, дает лучший компромисс между положительными эффектами и отрицательными побочными эффектами. [ 26 ]
Дофаминергические препараты, которые используются для лечения болезни Паркинсона, иногда связаны с развитием синдрома дизрегуляции дофамина , что включает в себя чрезмерное использование дофаминергических препаратов и индуцированного лекарством в природном вознаграждении, таких как азартные игры и сексуальная активность. [ 101 ] [ 102 ] Последнее поведение похоже на те, которые наблюдаются у людей с поведенческой зависимостью . [ 101 ]
Наркомания и психостимуляторы
[ редактировать ]
Кокаин , замещенные амфетамины (включая метамфетамин ), Adderall , метилфенидат (продаются как риталин или концерт ), а другие психостимуляторы оказывают свои эффекты, прежде всего, или частично, увеличивая уровень дофамина в мозге с помощью различных механизмов. [ 103 ] Кокаин и метилфенидат являются блокаторами дофамина или ингибиторам обратного захвата ; [ 104 ] Они не совпадают, ингибируют обратное захват дофамина, что приводит к повышению концентрации дофамина в синаптической расщелине. [ 105 ] [ 106 ] : 54–58 Как и кокаин, замещенные амфетамины и амфетамин также увеличивают концентрацию дофамина в синаптической расщелине , но с помощью различных механизмов. [ 36 ] [ 106 ] : 147–150
Эффекты психостимулянтов включают увеличение частоты сердечных сокращений, температуру тела и потоотделения; Улучшения в бдительности, внимании и выносливости; Увеличение удовольствия, производимого за счет вознаграждения событий; но в более высоких дозах возбуждение, беспокойство или даже потеря контакта с реальностью . [ 103 ] Препараты в этой группе могут иметь высокий потенциал зависимости из-за их активирующих эффектов на опосредованную дофамином систему вознаграждения в мозге. [ 103 ] Однако некоторые также могут быть полезны в более низких дозах для лечения дефицита внимания гиперактивности (СДВГ) и нарколепсии . [ 107 ] [ 108 ] Важным дифференцирующим фактором является начало и продолжительность действия. [ 103 ] Кокаин может вступить в силу в считанные секунды, если он вводится или вдыхается в свободной базовой форме; Эффекты длится от 5 до 90 минут. [ 109 ] Это быстрое и краткое действие делает его эффекты легко воспринимаемыми и, следовательно, дает ему высокий потенциал зависимости. [ 103 ] Метилфенидат, взятый в форме таблетки, напротив, может занять два часа, чтобы достичь пиковых уровней в крови, [ 107 ] И в зависимости от формулировки эффекты могут длиться до 12 часов. [ 110 ] Эти более длительные действия имеют преимущество в снижении потенциала для злоупотреблений и улучшению приверженности лечению с использованием более удобных схем дозировки. [ 111 ]

Разнообразные привыкающие препараты вызывают увеличение активности дофамина, связанной с вознаграждением. [ 103 ] Стимуляторы, такие как никотин , кокаин и метамфетамин, способствуют повышению уровня дофамина, которые, по -видимому, являются основным фактором, вызывающей зависимость. Для других привыкающих препаратов, таких как опиоидный героин, повышенный уровень дофамина в системе вознаграждения может играть лишь незначительную роль в зависимости. [ 112 ] Когда люди, зависимые от стимуляторов, проходят через снятие средств, они не испытывают физические страдания, связанные с отменой алкоголя или отменой из опиатов; Вместо этого они испытывают тягу, интенсивное стремление к препарату, характеризующемуся раздражительностью, беспокойством и другими симптомами возбуждения, [ 113 ] вызвано психологической зависимостью .
Система дофамина играет решающую роль в нескольких аспектах зависимости. На самой ранней стадии генетические различия, которые изменяют экспрессию рецепторов дофамина в мозге, могут предсказать, найдет ли человек стимуляторы привлекательными или неприятными. [ 114 ] Потребление стимуляторов вызывает повышение уровня дофамина мозга, которые длится от нескольких минут до часов. [ 103 ] Наконец, хроническое повышение дофамина, которое поставляется с повторяющимися высокими дозами стимулятора, вызывает широкий набор структурных изменений в мозге, которые ответственны за поведенческие аномалии, которые характеризуют зависимость. [ 115 ] Лечение стимуляторной зависимости очень сложно, потому что даже если потребление прекращается, тяга, которая сопровождается психологическим уходом. [ 113 ] Даже когда тяга, кажется, вымерла, она может вновь возникнуть, когда столкнутся со стимулами, которые связаны с препаратом, такими как друзья, местоположения и ситуации. [ 113 ] Ассоциационные сети в мозге значительно взаимосвязаны. [ 116 ]
Психоз и антипсихотические препараты
[ редактировать ]Психиатры в начале 1950 -х годов обнаружили, что класс лекарств, известных как типичные антипсихотики (также известные как основные транквилизаторы ), часто эффективны при снижении психотических симптомов шизофрении. [ 117 ] Введение первого широко используемого антипсихотического, хлорпромазина (торазин), в 1950 -х годах, привело к высвобождению многих пациентов с шизофренией из учреждений в последующие годы. [ 117 ] К 1970 -м годам исследователи понимали, что эти типичные антипсихотики работали в качестве антагонистов на рецепторах D 2 . [ 117 ] [ 118 ] Эта реализация привела к так называемой гипотезе дофамина о шизофрении , которая постулирует, что шизофрения в значительной степени вызвана гиперактивностью систем дофамина головного мозга. [ 119 ] Гипотеза допамина вызвала дополнительную поддержку от наблюдения, что психотические симптомы часто усиливались стимуляторами, повышающими дофамин, такие как метамфетамин, и что эти препараты также могут вызывать психоз у здоровых людей, если они принимаются в достаточно больших дозах. [ 119 ] В последующие десятилетия были разработаны другие атипичные антипсихотики , у которых было меньше серьезных побочных эффектов. [ 117 ] Многие из этих новых препаратов не действуют непосредственно на дофаминовые рецепторы, но вместо этого вызывают изменения в дофаминовой активности косвенно. [ 120 ] Эти препараты также использовались для лечения других психозов. [ 117 ] Антипсихотические препараты оказывают широко подавляющее влияние на большинство типов активного поведения и, в частности, снижают бредовое и возбужденное поведение, характерное для явного психоза. [ 118 ]
Однако более поздние наблюдения заставили гипотезу дофамина потерять популярность, по крайней мере, в его простой оригинальной форме. [ 119 ] Во -первых, пациенты с шизофренией, как правило, не демонстрируют измеримо повышенного уровня активности дофамина головного мозга. [ 119 ] Несмотря на это, многие психиатры и нейробиологи продолжают верить, что шизофрения включает в себя какую -то дисфункцию дофаминовой системы. [ 117 ] Однако, поскольку «гипотеза дофамина» развивалась с течением времени, виды дисфункций, которые она постулаты, как правило, становятся все более тонкими и сложными. [ 117 ]
Психофармаколог Стивен М. Шталь предположил в обзоре 2018 года, что во многих случаях психоза, включая шизофрению, три взаимосвязанные сети, основанные на дофамине, серотонине и глутамате - каждая из них самостоятельно или в различных комбинациях - способствовал чрезмерному экстриации допамина D 2. Рецепторы в вентральном стриатуме . [ 121 ]
Сэндрант дефицита внимания гиперактивности
[ редактировать ]Измененная нейротрансмиссия дофамина вовлечена в синдром гиперактивности дефицита внимания (СДВГ), условие, связанное с нарушением когнитивного контроля , в свою очередь, что приводит к проблемам с регулированием внимания ( контроль внимания ), ингибированию поведения ( ингибирующее контроль ) и забыванию или отсутствующим деталям ( работа Память ), среди других проблем. [ 122 ] Существуют генетические связи между дофаминовыми рецепторами, транспортером дофамина и СДВГ, в дополнение к связям с другими нейротрансмиттерными рецепторами и транспортерами. [ 123 ] Наиболее важная связь между дофамином и СДВГ включает препараты, которые используются для лечения СДВГ. [ 124 ] Некоторые из наиболее эффективных терапевтических агентов для СДВГ являются психостимулянтами, такими как метилфенидат (риталин, концерт) и амфетамин (Evekeo, Adderall, Dexedrine), препараты, которые увеличивают как уровни дофамина, так и норэпинефрина в мозге. [ 124 ] Клинические эффекты этих психостимулянтов при лечении СДВГ опосредуются посредством косвенной активации дофамина и норэпинефриновых рецепторов, в частности, дофаминового рецептора D 1 и адренорецептора α 2 , в префронтальной коре. [ 122 ] [ 125 ] [ 126 ]
Боль
[ редактировать ]Дофамин играет роль в обработке боли на нескольких уровнях центральной нервной системы, включая спинной мозг, периакедуктальный серый , таламус , базальные ганглии и поясной кору . [ 127 ] Снижение уровня дофамина была связана с болезненными симптомами, которые часто встречаются при болезни Паркинсона. [ 127 ] Аномалии в дофаминергической нейротрансмиссии также возникают в нескольких болезненных клинических состояниях, включая синдром горящего рта , фибромиалгию и синдром беспокойных ног. [ 127 ]
Тошнота
[ редактировать ]Тошнота и рвота в значительной степени определяются активностью в области постреми в медуле , ствола мозга в области, известной как зона триггера хеморецептора . [ 128 ] Эта область содержит большую популяцию дофаминовых рецепторов типа D 2 . [ 128 ] Следовательно, препараты, которые активируют рецепторы D 2, имеют высокий потенциал, вызывающий тошноту. [ 128 ] Эта группа включает в себя некоторые лекарства, которые вводятся для болезни Паркинсона, а также другие агонисты дофамина, такие как апоморфин . [ 129 ] В некоторых случаях антагонисты D 2 -рецепторов, такие как метоклопрамид, полезны в качестве анти -наезе . [ 128 ]
Сравнительная биология и эволюция
[ редактировать ]Микроорганизмы
[ редактировать ]нет сообщений о дофаминах В археи , но он был обнаружен в некоторых типах бактерий и в простейших, называемых тетрагименами . [ 130 ] Возможно, что еще более важно, существуют типы бактерий, которые содержат гомологи всех ферментов, которые животные используют для синтеза дофамина. [ 131 ] Было предложено, чтобы животные вывели их оборудование, синтезирующее дофамин из бактерий, посредством горизонтального переноса генов , которые могли произойти относительно поздно в эволюционное время, возможно, в результате симбиотического включения бактерий в эукариотические клетки, которые породили митохондрии . [ 131 ]
Животные
[ редактировать ]Дофамин используется в качестве нейротрансмиттера у большинства многоклеточных животных. [ 132 ] В губках есть только один отчет о наличии дофамина, без указания его функции; [ 133 ] Тем не менее, дофамин сообщалось в нервных системах многих других радиально симметричных видов, включая Cnidarian Jellyfish , Hydra и некоторые кораллы . [ 134 ] Это датируется появлением дофамина в качестве нейротрансмиттера обратно к самому раннему появлению нервной системы, более 500 миллионов лет назад в кембрийский период. Дофамин функционирует как нейротрансмиттер у позвоночных , эхинодерм , членистоногих , моллюсков и нескольких типов червя . [ 135 ] [ 136 ]
В любом типе животного, которое было изучено, был замечен дофамин модифицирует двигательное поведение. [ 132 ] В модельном организме , нематоде Caenorhabditis elegans , он уменьшает локомоцию и увеличивает пищевые экспларирующие движения; В плоских червях он производит «винтоподобные» движения; В пиявках он тормозит плавание и способствует ползу. В широком диапазоне позвоночных дофамин оказывает «активирующее» влияние на переключение поведения и отбор ответов, сравнимый с его влиянием у млекопитающих. [ 132 ] [ 137 ]
Дофамин также постоянно показал роль в обучении вознаграждениям во всех группах животных. [ 132 ] Как и у всех позвоночных - беспозвоночные , такие как круглые черви , плоские черви , моллюски и общие фруктовые мухи , могут быть обучены повторять действие, если оно последовательно сопровождается повышением уровней дофамина. [ 132 ] В плодовых мухах различные элементы для обучения вознаграждению предполагают модульную структуру для системы обработки вознаграждений насекомых, которая широко параллельна у млекопитающего. [ 138 ] Например, дофамин регулирует краткосрочное и долгосрочное обучение у обезьян; [ 139 ] У плодовых мух различные группы дофаминовых нейронов опосредуют сигналы вознаграждения для краткосрочных и долгосрочных воспоминаний. [ 140 ]
Давно считалось, что членистоногие были исключением из этого, когда дофамин рассматривается как неблагоприятный эффект. Награда была замечена, вместо этого было опосредовано октопамином , нейротрансмиттером, тесно связанным с норэпинефрином. [ 141 ] Более поздние исследования, однако, показали, что дофамин играет роль в обучении вознаграждениям в фруктовых мухах. Также было обнаружено, что полезный эффект октопамина связан с активированием набора дофаминергических нейронов, ранее не имеющих доступа к исследованию. [ 141 ]
Растения
[ редактировать ]
Многие растения, в том числе разнообразные пищевые растения, синтезируют дофамин в различной степени. [ 142 ] Самые высокие концентрации наблюдались в бананах - плодовая мякоть красных и желтых бананов содержит дофамин на уровне от 40 до 50 частей на миллион по весу. [ 142 ] Картофель, авокадо, брокколи и брюссельские капусты также могут содержать дофамин на уровне 1 части на миллион или более; Апельсины, помидоры, шпинат, бобы и другие растения содержат измеримые концентрации менее 1 части на миллион. [ 142 ] Дофамин в растениях синтезируется из аминокислотного тирозина, биохимическими механизмами, сходными с теми, которые используют животные. [ 142 ] Его можно метаболизировать различными способами, создавая меланин и различные алкалоиды в качестве побочных продуктов. [ 142 ] Функции катехоламинов растений не были четко установлены, но есть доказательства того, что они играют роль в ответе на стрессоры, такие как бактериальная инфекция, действуют в качестве факторов, способствующих росту, в некоторых ситуациях и модифицируют способ метаболизированы сахар. Рецепторы, которые опосредуют эти действия, еще не были идентифицированы, и не имеют внутриклеточных механизмов, которые они активируют. [ 142 ]
Дофамин, потребляемый в пище, не может действовать в мозг, потому что он не может преодолеть барьер кровь -мранка. [ 26 ] Тем не менее, существует также множество растений, которые содержат L-DOPA, метаболический предшественник дофамина. [ 143 ] Самые высокие концентрации обнаружены в листьях и стручках бобов растений рода слизи , особенно в мукуне pruriens (бархатные бобы), которые использовались в качестве источника для L-DOPA в качестве препарата. [ 144 ] Другим растением, содержащим значительное количество L-DOPA, является Vicia Faba , растение, которое производит фасоль Fava (также известную как «широкие бобы»). Однако уровень L-DOPA в бобах намного ниже, чем в оболочках стручков и других частях растения. [ 145 ] Семена деревьев кассии и баухинии также содержат значительные количества L-DOPA. [ 143 ]
У вида морских зеленых водорослей Ulvaria Obscura , основного компонента некоторых цветов водорослей , дофамин присутствует в очень высоких концентрациях, оценивается в 4,4% от сухого веса. Существуют доказательства того, что этот дофамин функционирует как защита от травоядных животных , снижая потребление улитками и изоподами . [ 146 ]
В качестве предшественника меланина
[ редактировать ]
Меланины-это семейство темно-пигментированных веществ, обнаруженных в широком спектре организмов. [ 147 ] Химически они тесно связаны с дофамином, и существует тип меланина, известный как дофамин-меланин , который может быть синтезирован путем окисления дофамина через фермент тирозиназу . [ 147 ] Меланин, который темнеет человеческая кожа, не относится к такому типу: он синтезируется путем, который использует L-DOPA в качестве предшественника, но не дофамина. [ 147 ] Тем не менее, есть убедительные доказательства того, что нейромеланин , который придает темный цвет субстанции мозга, по крайней мере частично, является дофамин-меланином. [ 148 ]
Меланин, полученный из дофамина, вероятно, также появляется, по крайней мере, в некоторых других биологических системах. Некоторые из дофамина в растениях, вероятно, будут использоваться в качестве предшественника дофамина-меланин. [ 149 ] Считается, что сложные узоры, которые появляются на крыльях бабочки, а также черно-белые полосы на тела личинок насекомых, также вызваны пространственно структурированными накоплениями дофамина-меланина. [ 150 ]
История и развитие
[ редактировать ]Допамин был впервые синтезирован в 1910 году Джорджем Баргером и Джеймсом Эвенсом в Wellcome Laboratories в Лондоне, Англия [ 151 ] и впервые идентифицировано в человеческом мозге Кэтрин Монтагю в 1957 году. Он был назван дофамином, потому что это моноамин , предшественник которого в синтезе Баргера-Эвенса составляет 3,4- d iHydr O Xy P Henyl A Lanine (Levodopa или L-Dopa) Полем Функция дофамина как нейротрансмиттера была впервые признана в 1958 году Арвидом Карлссоном и Нильс-оке Хилларп в Лаборатории химической фармакологии Национального института сердца Швеции . [ 152 ] Карлссон был удостоен Нобелевской премии 2000 года по физиологии или медицине за то, что дофамин является не только предшественником норэпинефрина (норадреналина) и адреналина (адреналин), но и сам по себе является нейтрансантером. [ 153 ]
Полидопамин
[ редактировать ]Исследования, мотивированные адгезивными полифенольными белками у мидий, привели к открытию в 2007 году, что широкий спектр материалов, если они помещены в раствор дофамина при слегка базовом pH , будут покрыты слоем полимеризованного дофамина, часто называемого полидопамином . [ 154 ] [ 155 ] Этот полимеризованный дофамин образуется с помощью спонтанной реакции окисления и формально является типом меланина. [ 156 ] Кроме того, самополимеризация дофамина может использоваться для модуляции механических свойств гелей на основе пептидов. [ 157 ] Синтез полидопамина обычно включает реакцию гидрохлорида дофамина с Трис в качестве основания в воде. Структура полидопамина неизвестна. [ 155 ]
Полидопаминовые покрытия могут образовываться на объектах, варьирующихся в размере от наночастиц до больших поверхностей. [ 156 ] Полидопаминовые слои обладают химическими свойствами, которые могут быть чрезвычайно полезными, и многочисленные исследования изучали их возможные применения. [ 156 ] На самом простом уровне их можно использовать для защиты от повреждения светом или для формирования капсул для доставки лекарств. [ 156 ] На более сложном уровне их адгезивные свойства могут сделать их полезными в качестве субстратов для биосенсоров или других биологически активных макромолекул. [ 156 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Cruickshank L, Kennedy AR, Shankland N (2013). «Вход CSD Тирзакс: 5- (2-аммониоэтил) -2-гидроксифенолат, дофамин» . Кембриджская структурная база данных: конструкции доступа . Кембриджский кристаллографический центр обработки данных . doi : 10.5517/cc10m9nl .
- ^ Cruickshank L, Kennedy AR, Shankland N (2013). «Таутомерные и ионизационные формы дофамина и тирамина в твердом состоянии». J. Mol. Структура 1051 : 132–36. BIBCODE : 2013JST1051..132C . doi : 10.1016/j.molstruc.2013.08.002 .
- ^ Jump up to: а беременный в дюймовый «Дофамин: биологическая активность» . Руководство IUPHAR/BPS по фармакологии . Международный союз базовой и клинической фармакологии . Получено 29 января 2016 года .
- ^ Берридж К.С. (апрель 2007 г.). «Дебаты о роли дофамина в вознаграждении: дело о поощрительной значимости». Психофармакология . 191 (3): 391–431. doi : 10.1007/s00213-006-0578-x . PMID 17072591 . S2CID 468204 .
- ^ Jump up to: а беременный Мудрый Р.А., Роббл М.А. (январь 2020 г.). «Дофамин и зависимость» . Ежегодный обзор психологии . 71 (1): 79–106. doi : 10.1146/annurev-psych-010418-103337 . PMID 31905114 . S2CID 210043316 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Malenka RC, Nestler EJ, Hyman SE (2009). Sydor A, Brown Ry (Eds.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. С. 147–48, 366–67, 375–76. ISBN 978-0-07-148127-4 .
- ^ Балики М.Н., Мансур А., Бария А.Т., Хуан Л., Бергер С.Е., Филдс Х.Л., Апкирян Ав (октябрь 2013 г.). «Размещение человека укупается в предполагаемое ядро и раковину диссоциирует кодирование ценностей для вознаграждения и боли» . Журнал нейробиологии . 33 (41): 16383–93. doi : 10.1523/jneurosci.1731-13.2013 . PMC 3792469 . PMID 24107968 .
- ^ Jump up to: а беременный Венцель Дж. М., Раушер Н.А., Черт Дж. Ф., Олесон Э.Б. (январь 2015 г.). «Роль фазического высвобождения дофамина в ядре, наполненном кодированием отвращения: обзор нейрохимической литературы» . ACS Химическая нейробиология . 6 (1): 16–26. doi : 10.1021/cn500255p . PMC 5820768 . PMID 25491156 .
Таким образом, вызванные страхом стимулы способны дифференциально изменять передачу фазовой дофамина в субрегионах NACC. Авторы предполагают, что наблюдаемое улучшение в дофамине NACC Shell, вероятно, отражает общую мотивационную значимость, возможно, из-за облегчения от CS-индуцированного состояния страха, когда США (шок) не доставляется. Это рассуждение поддерживается докладом Budygin и Colleags 112 Показывая, что у анестезированных крыс завершение закрепления хвоста приводит к увеличению высвобождения дофамина в оболочке.
- ^ Puglisi-Allegra S, Ventura R (июнь 2012 г.). «Префронтальная/прикачная катехоламиновая система обрабатывает высокую мотивационную значимость» . Передний. Поведение Нейроски . 6 : 31. doi : 10.3389/fnbeh.2012.00031 . PMC 3384081 . PMID 22754514 .
- ^ Moncrieff J (2008). Миф о химическом лечении. Критика психиатрического лечения наркотиков . Бейсингсток, Великобритания: Palgrave Macmillan. ISBN 978-0-230-57432-8 .
- ^ Volkow ND, Wang GJ, Kollins SH, Wigal TL, Newcorn JH, Telang F, Fowler JS, Zhu W, Logan J, Ma Y, Pradhan K, Wong C, Swanson JM (сентябрь 2009 г.). «Оценка пути вознаграждения дофамина при СДВГ: клинические последствия» . Джама . 302 (10): 1084–91. doi : 10.1001/Jama.2009.1308 . PMC 2958516 . PMID 19738093 .
- ^ «Инфузия дофамина» (PDF) . Получено 13 октября 2023 года .
- ^ Jump up to: а беременный «Шок и гипотония при препаратах новорожденных: альфа/бета -адренергические агонисты, вазодилататоры, инотропные агенты, расширители объема, антибиотики, другие» . emedicine.medscape.com . Получено 13 октября 2023 года .
- ^ «Дофамин» . Pubchem . Получено 21 сентября 2015 года .
- ^ "Катехоламин" . Британская . Получено 21 сентября 2015 года .
- ^ «Фенилэтиламин» . Chemicalland21.com . Получено 21 сентября 2015 года .
- ^ Jump up to: а беременный в дюймовый и фон Картер Дж., Джонсон Дж.Х., Бааске Д.М. (1982). «Дофамин гидрохлорид». Аналитические профили лекарственных веществ . 11 : 257–72. doi : 10.1016/s0099-5428 (08) 60266-x . ISBN 978-0122608117 .
- ^ "Спецификационный лист" . www.sigmaaldrich.com . Получено 13 сентября 2019 года .
- ^ Jump up to: а беременный в Broadley KJ (март 2010 г.). «Сосудистые эффекты следов аминов и амфетаминов». Фармакология и терапия . 125 (3): 363–375. doi : 10.1016/j.pharmthera.2009.11.005 . PMID 19948186 .
- ^ Jump up to: а беременный в Lindemann L, Hoener MC (май 2005 г.). «Ренессанс в Trace Amines, вдохновленный новой семейством GPCR». Тенденции в фармакологических науках . 26 (5): 274–281. doi : 10.1016/j.tips.2005.03.007 . PMID 15860375 .
- ^ Jump up to: а беременный в дюймовый Wang X, Li J, Dong G, Yue J (февраль 2014 г.). «Эндогенные субстраты мозга CYP2D». Европейский журнал фармакологии . 724 : 211–218. doi : 10.1016/j.ejphar.2013.12.025 . PMID 24374199 .
- ^ Jump up to: а беременный в дюймовый и фон Симан П. (2009). «Глава 1: Исторический обзор: Введение в дофаминовые рецепторы». В Neve K (ред.). Рецепторы дофамина . Спрингер. С. 1–22. ISBN 978-1-60327-333-6 .
- ^ «EC 1.14.16.2-тирозин 3-монооксигеназы (Homo sapiens)» . Бренда . Технический университет Брауншвейга. Июль 2016 года . Получено 7 октября 2016 года .
Субстраты: L-фенилаланин + тетрагидробиоптер + O2
Продукт: L-тирозин + 3-гидроксифенилаланин [(ака м-тирозин)] + дигидроптеридин + H2O
Организм: Homo Sapiens
Диаграмма реакции - ^ «EC 4.1.1.28-ароматическая L-аминокислотная декарбоксилаза (Homo sapiens)» . Бренда . Technische Universität Braunschweig. Июль 2016 года . Получено 7 октября 2016 года .
Субстрат: м-тирозин
Продукт: m-tyramine + co2
Организм: Homo Sapiens
Диаграмма реакции - ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Musacchio JM (2013). «Глава 1: Ферменты, участвующие в биосинтезе и деградации катехоламинов». В Iverson L (ред.). Биохимия биогенных аминов . Спрингер. С. 1–35. ISBN 978-1-4684-3171-1 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Национальный сотрудничающий центр хронических состояний, изд. (2006). «Симптоматическая фармакологическая терапия при болезни Паркинсона» . Болезнь Паркинсона . Лондон: Королевский колледж врачей. С. 59–100. ISBN 978-1-86016-283-1 Полем Архивировано из оригинала 24 сентября 2010 года . Получено 24 сентября 2015 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Eisenhofer G, Kopin IJ, Goldstein DS (сентябрь 2004 г.). «Катехоламиновый метаболизм: современный взгляд с последствиями для физиологии и медицины». Фармакологические обзоры . 56 (3): 331–49. doi : 10.1124/pr.56.3.1 . PMID 15317907 . S2CID 12825309 .
- ^ Zahoor I, Shafi A, Haq E (декабрь 2018 г.). «Фармакологическое лечение болезни Паркинсона: Рисунок 1: [Метаболический путь синтеза дофамина ...]» . В Stoker TB, Greenland JC (Eds.). Болезнь Паркинсона: патогенез и клинические аспекты [Интернет] . Брисбен (AU): Кодовые публикации.
- ^ Амин Ф., Дэвидсон М., Дэвис К.Л. (1992). «Измерение гомованиллической кислоты в клинических исследованиях: обзор методологии» . Бюллетень шизофрении . 18 (1): 123–48. doi : 10.1093/schbul/18.1.123 . PMID 1553492 .
- ^ Амин Ф., Дэвидсон М., Кан Р.С., Шмейдлер Дж., Стерн Р., Нотт П.Дж., Аптер С. (1995). «Оценка центрального дофаминергического индекса плазменного HVA при шизофрении» . Бюллетень шизофрении . 21 (1): 53–66. doi : 10.1093/schbul/21.1.53 . PMID 7770741 .
- ^ Sulzer D, Zecca L (февраль 2000 г.). «Интранерональный синтез дофамина-хинона: обзор». Исследование нейротоксичности . 1 (3): 181–95. doi : 10.1007/bf03033289 . PMID 12835101 . S2CID 21892355 .
- ^ Миядзаки I, Асанума М (июнь 2008 г.). «Дофаминергический нейрон-специфический окислительный стресс, вызванный самим дофамином» (PDF) . Acta Medica Okmama . 62 (3): 141–50. doi : 10.18926/amo/30942 . PMID 18596830 .
- ^ Jump up to: а беременный в дюймовый и Grandy DK, Miller GM, Li JX (февраль 2016 г.). « Taargeting Addiction» - Alamo Bears Свидетель на другую революцию: обзор пленарного симпозиума по поведению, биологии и химии 2015 года » . Наркотики и алкогольная зависимость . 159 : 9–16. doi : 10.1016/j.drugalcdep.2015.11.014 . PMC 4724540 . PMID 26644139 .
TAAR1 является рецептором с высоким аффингом для метамфетамина/AMPH и DA
- ^ Jump up to: а беременный в дюймовый Romanelli RJ, Williams JT, Neve Ka (2009). «Глава 6: передача сигналов рецептора дофамина: внутриклеточные пути к поведению». В Neve Ka (ред.). Рецепторы дофамина . Спрингер. С. 137–74. ISBN 978-1-60327-333-6 .
- ^ Jump up to: а беременный Эйден Л.Е., Шефер М.К., Вейхе Е., Шюц Б. (февраль 2004 г.). «Семейство переносчиков везикулярных амина (SLC18): антипортеры амина/протонов, необходимые для везикулярного накопления и регулируемой экзоцитотической секреции моноаминов и ацетилхолина». Pflügers Archiv . 447 (5): 636–40. doi : 10.1007/s00424-003-1100-5 . PMID 12827358 . S2CID 20764857 .
- ^ Jump up to: а беременный Miller GM (январь 2011 г.). «Новая роль связанного с амином рецептора 1 в функциональной регуляции моноаминовых транспортеров и дофаминергической активности» . Журнал нейрохимии . 116 (2): 164–76. doi : 10.1111/j.1471-4159.2010.07109.x . PMC 3005101 . PMID 21073468 .
- ^ Jump up to: а беременный Beaulieu JM, Gainetdinov RR (март 2011 г.). «Физиология, передача сигналов и фармакология дофаминовых рецепторов». Фармакологические обзоры . 63 (1): 182–217. doi : 10.1124/pr.110.002642 . PMID 21303898 . S2CID 2545878 .
- ^ Torres GE, Gainetdinov RR, Caron MG (январь 2003 г.). «Перевороты плазматической мембраны: структура, регуляция и функция». Природные обзоры. Нейробиология . 4 (1): 13–25. doi : 10.1038/nrn1008 . PMID 12511858 . S2CID 21545649 .
- ^ Jump up to: а беременный в дюймовый Rice Me, Patel JC, Cragg SJ (декабрь 2011 г.). «Выпуск дофамина в базальных ганглиях» . Нейробиология . 198 : 112–37. doi : 10.1016/j.neuroscience.2011.08.066 . PMC 3357127 . PMID 21939738 .
- ^ Schultz W (2007). «Множественные функции дофамина на разных временных курсах». Ежегодный обзор нейробиологии . 30 : 259–88. doi : 10.1146/annurev.neuro.28.061604.135722 . PMID 17600522 . S2CID 13503219 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Björklund A, Dunnett SB (май 2007). «Системы дофаминовых нейронов в мозге: обновление». Тенденции в нейронауках . 30 (5): 194–202. doi : 10.1016/j.tins.2007.03.006 . PMID 17408759 . S2CID 14239716 .
- ^ Jump up to: а беременный Dahlstroem A, Fuxe K (1964). «Свидетельство о существовании моноаминосодержащих нейронов в центральной нервной системе. I. Демонстрация моноаминов в клеточных телах нейронов ствола головного мозга». Acta Physiologica Scandinavica. Дополнение . 232 (Suppl): 1–55. PMID 14229500 .
- ^ Jump up to: а беременный в дюймовый Malenka RC, Nestler EJ, Hyman SE (2009). «Глава 6: широко выступающие системы: моноамины, ацетилхолин и орексин». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. С. 147–48, 154–57. ISBN 978-0-07-148127-4 .
- ^ Кристина К.В., Аминофф М.Дж. (сентябрь 2004 г.). «Клиническая дифференциация паркинсонских синдромов: прогностическая и терапевтическая значимость». Американский журнал медицины . 117 (6): 412–19. doi : 10.1016/j.amjmed.2004.03.032 . PMID 15380498 .
- ^ Fadok JP, Dickerson TM, Palmiter Rd (сентябрь 2009 г.). «Дофамин необходим для зависимой от сигнала страха» . Журнал нейробиологии . 29 (36): 11089–97. doi : 10.1523/jneurosci.1616-09.2009 . PMC 2759996 . PMID 19741115 .
- ^ Тан В., Кохуби О., Кинчер М., Шнеггенбургер Р. (апрель 2020 г.). «Проекция VTA для базальной миндалины дофамина способствует сигналу значительных соматосенсорных событий во время обучения страха» . Журнал нейробиологии . 40 (20): JN-RM-1796-19. doi : 10.1523/jneurosci.1796-19.2020 . PMC 7219297 . PMID 32277045 .
- ^ Jo YS, Heymann G, Zweifel LS (ноябрь 2018 г.). «Дофаминовые нейроны отражают неопределенность в обобщении страха» . Нейрон . 100 (4): 916–925.e3. doi : 10.1016/j.neuron.2018.09.028 . PMC 6226002 . PMID 30318411 .
- ^ Jump up to: а беременный Паулюс В., Шомбург Эд (июнь 2006 г.). «Дофамин и спинной мозг при синдроме беспокойных ног: выявляет ли физиология спинного мозга основание для увеличения?». Обзоры медицины сна . 10 (3): 185–96. doi : 10.1016/j.smrv.2006.01.004 . PMID 16762808 .
- ^ Jump up to: а беременный в дюймовый и Бен-Джонатан Н., Хнаско Р (декабрь 2001 г.). «Дофамин как ингибитор пролактина (PRL)» . Эндокринные обзоры . 22 (6): 724–63. doi : 10.1210/er.22.6.724 . PMID 11739329 .
- ^ Jump up to: а беременный в дюймовый Витковский П (январь 2004 г.). «Функция дофамина и сетчатки» . Documenta Ophthalmologica. Достижения в офтальмологии . 108 (1): 17–40. doi : 10.1023/b: doop.0000019487.88486.0a . PMID 15104164 . S2CID 10354133 .
- ^ Jump up to: а беременный Исправить JD (2008). «Базальные ганглии и стриатальная моторная система». Нейроанатомия (Series Series) (4 -е изд.). Балтимор: Wulters Kluwer & Lippincott Williams & Wilkins. С. 274–81. ISBN 978-0-7817-7245-7 .
- ^ Jump up to: а беременный в дюймовый и фон Чакраварти В.С., Джозеф Д., Бапи Р.С. (сентябрь 2010). «Что делают базальные ганглии? Перспектива моделирования» . Биологическая кибернетика . 103 (3): 237–53. doi : 10.1007/s00422-010-0401-y . PMID 20644953 . S2CID 853119 .
- ^ Jump up to: а беременный в дюймовый Floresco SB (январь 2015). «Прилегающее ядро: интерфейс между познанием, эмоциями и действием» . Ежегодный обзор психологии . 66 : 25–52. doi : 10.1146/annurev-psych-010213-115159 . PMID 25251489 . S2CID 28268183 .
- ^ Jump up to: а беременный Balleine BW, Dezfouli A, Ito M, Doya K (2015). «Иерархический контроль над целенаправленным действием в сети коры-базальных ганглиев». Текущее мнение в поведенческих науках . 5 : 1–7. doi : 10.1016/j.cobeha.2015.06.001 . S2CID 53148662 .
- ^ Jump up to: а беременный в Jankovic J (апрель 2008 г.). «Болезнь Паркинсона: клинические особенности и диагноз» . Журнал неврологии, нейрохирургии и психиатрии . 79 (4): 368–76. doi : 10.1136/jnnp.2007.131045 . PMID 18344392 .
- ^ Pattij T, Vanderschuren LJ (апрель 2008 г.). «Нейрофармакология импульсивного поведения» . Тенденции в фармакологических науках . 29 (4): 192–99. doi : 10.1016/j.tips.2008.01.002 . PMID 18304658 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Schultz W (июль 2015 г.). «Нейрональные вознаграждения и сигналы принятия решений: от теорий до данных» . Физиологические обзоры . 95 (3): 853–951. doi : 10.1152/physrev.00023.2014 . PMC 4491543 . PMID 26109341 .
- ^ Jump up to: а беременный в Робинсон Т.Е., Берридж К.С. (1993). «Нейронная основа жалки наркотиков: теория стимулирующей сенсибилизации зависимости». Исследование мозга. Обзоры исследований мозга . 18 (3): 247–91. doi : 10.1016/0165-0173 (93) 90013-p . HDL : 2027.42/30601 . PMID 8401595 . S2CID 13471436 .
- ^ Wright JS, Panksepp J (2012). «Эволюционная структура для понимания кормления, желания и желания: нейропсихология системы поиска» . Нейропсихоанализ . 14 (1): 5–39. doi : 10.1080/15294145.2012.10773683 . S2CID 145747459 . Получено 24 сентября 2015 года .
- ^ Jump up to: а беременный в дюймовый и Берридж К.С., Робинсон Т.Е., Олдридж Дж.В. (февраль 2009 г.). «Расширение компонентов вознаграждения:« Любие »,« Желание »и обучение» . Современное мнение о фармакологии . 9 (1): 65–73. doi : 10.1016/j.coph.2008.12.014 . PMC 2756052 . PMID 19162544 .
- ^ Montague PR, Dayan P, Sejnowski TJ (март 1996 г.). «Структура для мезоэфальных дофаминовых систем на основе прогнозирующего обучения иббиян» . Журнал нейробиологии . 16 (5): 1936–47. doi : 10.1523/jneurosci.16-05-01936.1996 . PMC 6578666 . PMID 8774460 .
- ^ Bromberg-Martin ES, Matsumoto M, Hikosaka O (декабрь 2010 г.). «Дофамин в мотивационном контроле: вознаграждение, отвращение и предупреждение» . Нейрон . 68 (5): 815–34. doi : 10.1016/j.neuron.2010.11.022 . PMC 3032992 . PMID 21144997 .
- ^ Ягер Л.М., Гарсия А.Ф., Вунш А.М., Фергюсон С.М. (август 2015 г.). «Входы и выходы стриатума: роль в наркомании» . Нейробиология . 301 : 529–41. doi : 10.1016/j.neuroscience.2015.06.033 . PMC 4523218 . PMID 26116518 .
- ^ Jump up to: а беременный MP Saddoris, Cacciapaglia F, Wightman RM, Carelli RM (август 2015 г.). «Дифференциальная динамика высвобождения дофамина в ядре Accumbens Core и Shell выявляет комплементарные сигналы для прогнозирования ошибок и стимулирующей мотивации» . Журнал нейробиологии . 35 (33): 11572–82. doi : 10.1523/jneurosci.2344-15.2015 . PMC 4540796 . PMID 26290234 .
- ^ Берридж К.С., Крингельбах М.Л. (май 2015). «Системы удовольствия в мозге» . Нейрон . 86 (3): 646–64. doi : 10.1016/j.neuron.2015.02.018 . PMC 4425246 . PMID 25950633 .
- ^ Jump up to: а беременный в Мудрый Р.А. (1996). «Привыкание препаратов и награды за стимуляцию мозга». Ежегодный обзор нейробиологии . 19 : 319–40. doi : 10.1146/annurev.ne.19.030196.001535 . PMID 8833446 .
- ^ Мудрый Р.А. (октябрь 2008 г.). «Дофамин и награда: гипотеза Анедонии 30 лет» . Исследование нейротоксичности . 14 (2–3): 169–83. doi : 10.1007/bf03033808 . PMC 3155128 . PMID 19073424 .
- ^ Arias-Carrión O, Pöppel E (2007). «Дофамин, обучение и поведение, обращаясь за наградами» . Acta Neurobiol Exp . 67 (4): 481–88. doi : 10.55782/ane-2007-1664 . PMID 18320725 .
- ^ Ikemoto S (ноябрь 2007 г.). «Схема вознаграждения дофамина: две проекционные системы от вентрального среднего мозга до ядра прикупаемого туберкулеза» . Обзоры исследований мозга . 56 (1): 27–78. doi : 10.1016/j.brainresrev.2007.05.004 . PMC 2134972 . PMID 17574681 .
- ^ Jump up to: а беременный Феррери Л., Мас-Херреро Е., Заторр Р.Дж., Рипеллес П., Гомес-Андрес А., Аликарт Х., Олив Г., Марко-Плалларес Дж., Антониджоан Р.М., Валле М., Риба Дж., Родригес-Форнеллс А (2019). «Дофамин модулирует вознаграждение, вызванное музыкой» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3793–98. Bibcode : 2019pnas..116.3793f . doi : 10.1073/pnas.1811878116 . PMC 6397525 . PMID 30670642 .
Прослушивание приятной музыки часто сопровождается измеримыми телесными реакциями, такими как гусиные удары или дрожь по позвоночнику, обычно называемую «озноб» или «Frissons». ... В целом, наши результаты прямо показали, что фармакологические вмешательства двунаправленно модулировали ответы вознаграждения, вызванные музыкой. В частности, мы обнаружили, что рисперидон нарушает способность участников испытывать музыкальное удовольствие, тогда как леводопа улучшила его. ... Здесь, напротив, изучение ответов на абстрактные награды у людей, мы показываем, что манипулирование дофаминергической передачей влияет как на удовольствие (то есть, количество времени, сообщающих о охлаждении, и эмоциональном возбуждении, измеряемом EDA), так и на мотивационные компоненты музыкального вознаграждения и эмоциональное возбуждение, измеренное EDA), так и на мотивационные компоненты музыкального вознаграждения и эмоциональное возбуждение (деньги, желающие потратить). Эти данные свидетельствуют о том, что дофаминергическая передача сигналов является не только условием не только для мотивационных ответов, как было показано с первичными и вторичными наградами, но и для гедонистических реакций на музыку. Этот результат подтверждает недавние результаты, показывающие, что дофамин также опосредует воспринимаемую приятность, достигнутую другими типами абстрактных вознаграждений (37), и вызовет предыдущие результаты на животных моделях на первичных вознаграждениях, таких как пища (42, 43).
- ^ Jump up to: а беременный Goupil L, Aucouturier JJ (февраль 2019 г.). «Музыкальное удовольствие и музыкальные эмоции» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3364–66. Bibcode : 2019pnas..116.3364G . doi : 10.1073/pnas.1900369116 . PMC 6397567 . PMID 30770455 .
В фармакологическом исследовании, опубликованном в PNAS, Ferreri et al. (1) Представьте доказательства, которые усиление или ингибирование передачи сигналов дофамина с использованием леводопы или рисперидона модулирует удовольствие при прослушивании музыки. ... в последнем залже, чтобы установить не только корреляционное, но и причинное значение дофамина в музыкальном удовольствии, авторы обратились к непосредственному манипулированию дофаминергической передачи сигналов в стриатуме, сначала применив возбуждающие и ингибирующие транскраниальные магнитные стимуляции над своими участниками ' Левая дорсолатеральная префронтальная кора, область, известная для модуляции стриатальной функции (5), и, наконец, в настоящем исследовании, путем введения фармацевтических агентов, способных изменить доступность синаптической дофамина (1), оба из которых повлияли на воспринимаемое удовольствие, физиологические показатели возбуждения, и денежное значение, назначенное музыке в прогнозируемом направлении. ... хотя вопрос о музыкальном выражении эмоций имеет долгую историю исследования, в том числе в PNAS (6), и психофизиологическая направление исследований 1990 -х годов уже установила, что музыкальное удовольствие может активировать вегетативную нервную систему (7), The) Авторы демонстрация значения системы вознаграждения в музыкальных эмоциях была принята как инаугурационное доказательство того, что это были верные эмоции, чьи исследования имеют полную легитимность, чтобы информировать нейробиологию наших повседневных когнитивных, социальных и аффективных функций (8). Кстати, эта линия работы, кульминацией которой является статья Ferreri et al. (1), правдоподобно сделал больше для привлечения финансирования исследований для области музыкальных наук, чем любой другой в этом сообществе.
Свидетельство Ferreri et al. (1) обеспечивает последнюю поддержку убедительной нейробиологической модели, в которой музыкальное удовольствие возникает из -за взаимодействия древних систем вознаграждения/оценки (стриатально -лимбик -паралимбическая) с более филогенетически продвинутыми системами восприятия/прогнозов (временному течению). - ^ Koepp MJ, Gunn RN, Lawrence AD, Cunningham VJ, Dagher A, Jones T, et al. (Май 1998). «Свидетельство для выпуска дофамина стриата во время видеоигры». Природа . 393 (6682): 266–268. Bibcode : 1998natur.393..266k . doi : 10.1038/30498 . PMID 9607763 . S2CID 205000565 .
- ^ Von Der Heiden JM, Braun B, Müller KW, Egloff B (2019). «Связь между видеоигры и психологическим функционированием» . Границы в психологии . 10 : 1731. DOI : 10.3389/fpsyg.2019.01731 . PMC 6676913 . PMID 31402891 .
- ^ Jump up to: а беременный Missale C, Nash SR, Robinson SW, Jaber M, Caron MG (январь 1998 г.). «Рецепторы дофамина: от структуры к функционированию» (PDF) . Физиологические обзоры . 78 (1): 189–225. doi : 10.1152/physrev.1998.78.1.189 . PMID 9457173 . S2CID 223462 . Архивировано из оригинала (PDF) 2 марта 2019 года.
- ^ Jump up to: а беременный Buttarelli FR, Fanciulli A, Pellicano C, Pontieri Fe (июнь 2011 г.). «Дофаминергическая система в лимфоцитах периферической крови: от физиологии до фармакологии и потенциальных применений к психоневрологическим расстройствам» . Текущая нейрофармакология . 9 (2): 278–88. doi : 10.2174/157015911795596612 . PMC 3131719 . PMID 22131937 .
- ^ Jump up to: а беременный Саркар С., Басу Б., Чакробота Д., Дасгупта П.С., Басу С (май 2010 г.). «Иммунорегуляторная роль дофамина: обновление» . Мозг, поведение и иммунитет . 24 (4): 525–28. doi : 10.1016/j.bbi.2009.10.015 . PMC 2856781 . PMID 19896530 .
- ^ Хуссейн Т., Локхандвала М.Ф. (февраль 2003 г.). «Почечные дофаминовые рецепторы и гипертония». Экспериментальная биология и медицина . 228 (2): 134–42. doi : 10.1177/153537020322800202 . PMID 12563019 . S2CID 10896819 .
- ^ Choi MR, Kouyoumdzian NM, Rukavina Mikusic NL, Kravetz MC, Rosón Mi, Rodríguez Fermepin M, Fernández BE (май 2015). «Почечная дофаминергическая система: патофизиологические последствия и клинические перспективы» . Мировой журнал нефрологии . 4 (2): 196–212. doi : 10.5527/wjn.v4.i2.196 . PMC 4419129 . PMID 25949933 .
- ^ Кэри Р.М. (сентябрь 2001 г.). «Лекция Теодора Купера: почечная дофаминовая система: паракринный регулятор гомеостаза натрия и артериального давления» . Гипертония . 38 (3): 297–302. doi : 10.1161/hy0901.096422 . PMID 11566894 .
- ^ Jump up to: а беременный в дюймовый и Rubí B, Maechler P (декабрь 2010 г.). «Minireview: новые роли периферического дофамина в отношении метаболического контроля и роста опухоли: давайте искать баланс» . Эндокринология . 151 (12): 5570–81. doi : 10.1210/en.2010-0745 . PMID 21047943 .
- ^ «Кто моделирует список основных лекарств» (PDF) . Всемирная организация здравоохранения . Октябрь 2013 г. Архивировал (PDF) с оригинала 10 февраля 2014 года . Получено 24 сентября 2015 года .
- ^ Noori S, Friedlich P, Seri I (2003). «Фармакологический обзор, регулируемый развитием сердечно -сосудистых, почечных и нейроэндокринных эффектов дофамина» . Neoreviews . 4 (10): E283 - E288. doi : 10.1542/neo.4-10-e283 . S2CID 71902752 . Получено 24 сентября 2015 года .
- ^ Jump up to: а беременный Bhatt-Mehta V, Nahata MC (1989). «Дофамин и добутамин при педиатрической терапии». Фармакотерапия . 9 (5): 303–14. doi : 10.1002/j.1875-9114.1989.tb04142.x . PMID 2682552 . S2CID 25614283 .
- ^ Jump up to: а беременный в Bronwen JB, Knights KM (2009). Фармакология для медицинских работников (2 -е изд.). Elsevier Australia. п. 192. ISBN 978-0-7295-3929-6 .
- ^ De Backer D, Biston P, Devriendt J, Madl C, Chochrad D, Aldecoa C, et al. (Март 2010 г.). «Сравнение дофамина и норэпинефрина при лечении шока» (PDF) . Новая Англия Журнал медицины . 362 (9): 779–89. doi : 10.1056/nejmoa0907118 . PMID 20200382 . S2CID 2208904 . Архивировано из оригинала (PDF) 28 февраля 2019 года.
- ^ Картик С., Лиссабон А. (2006). «Низкие дозы дофамина в отделении интенсивной терапии». Семинары в диализе . 19 (6): 465–71. doi : 10.1111/j.1525-139x.2006.00208.x . PMID 17150046 . S2CID 22538344 .
- ^ Моисей С. "дофамин" . Семейная практика ноутбука . Получено 1 февраля 2016 года .
- ^ Катритсис Д.Г., Герш Б.Дж., Камм А.Дж. (2013). Клиническая кардиология: текущие практические руководящие принципы . УП Оксфорд. ISBN 978-0-19-150851-6 Полем
Дофамин связывается с бета-1, бета-2, альфа-1 и дофаминергическими рецепторами
- ^ Льюис Р.Дж. (2004). Опасные свойства Саксорога промышленных материалов (11 -е изд.). Хобокен, Нью -Джерси: Wiley & Sons. п. 1552. ISBN 978-0-471-47662-7 .
- ^ Standaert DG, Walsh RR (2011). «Фармакология дофаминергической нейротрансмиссии». В Ташжяне А.Х., Армстронг Э.Дж., Голан де (ред.). Принципы фармакологии: патофизиологическая основа лекарственной терапии . Липпинкотт Уильямс и Уилкинс. С. 186–206. ISBN 978-1-4511-1805-6 .
- ^ Mobbs CV, HOF PR (2009). Справочник по нейробиологии старения . Амстердам: Elsevier/Academic Press. ISBN 978-0-12-374898-0 Полем OCLC 299710911 .
- ^ Ota M, Yasuno F, Ito H, Seki C, Nozaki S, Asada T, Suhara T (июль 2006 г.). «Возрастное снижение синтеза дофамина в живом человеческом мозге, измеренное по позитронной эмиссионной томографии с L- [Beta-11C] Dopa». Жизненные науки . 79 (8): 730–36. doi : 10.1016/j.lfs.2006.02.017 . PMID 16580023 .
- ^ Kaasinen V, Vilkman H, Hietala J, Någren K, Helenius H, Olsson H, Farde L, Rinne J (2000). «Возрастная потеря дофамина D2/D3 потери рецептора в экстрастриатских областях человеческого мозга». Нейробиология старения . 21 (5): 683–68. doi : 10.1016/s0197-4580 (00) 00149-4 . PMID 11016537 . S2CID 40871554 .
- ^ Ван Ю., Чан Г.Л., Холден Дж. Э., Добко Т., Мак Е., Шулцер М., Хузер Дж. М., Сноу Б.Дж., Рут Т.Дж., Кальн Д.Б., Стоссл А.Дж. (сентябрь 1998 г.). «Возрастное снижение рецепторов дофамина D1 в мозге человека: исследование домашних животных». Синапс . 30 (1): 56–61. doi : 10.1002/(sici) 1098-2396 (199809) 30: 1 <56 :: Aid-Syn7> 3.0.co; 2-j . PMID 9704881 . S2CID 31445572 .
- ^ Wong DF, Wagner HN, Dannals RF, Links JM, Frost JJ, Ravert HT, Wilson AA, Rosenbaum AE, Gjedde A, Douglass KH (декабрь 1984 г.). «Влияние возраста на рецепторы дофамина и серотонина, измеренные по позитронному томографии в живом человеческом мозге». Наука . 226 (4681): 1393–96. Bibcode : 1984sci ... 226.1393W . doi : 10.1126/science.6334363 . PMID 6334363 . S2CID 24278577 .
- ^ Jump up to: а беременный Wang E, Snyder SD (1998). Справочник по стареющему мозгу . Сан -Диего, Калифорния: академическая пресса. ISBN 978-0-12-734610-6 Полем OCLC 636693117 .
- ^ Dobryakova E, Genova HM, Deluca J, Wylie GR (12 марта 2015 г.). «Гипотеза дисбаланса дофамина о усталости при рассеянном склерозе и других неврологических расстройствах» . Границы в неврологии . 6 : 52. DOI : 10.3389/fneur.2015.00052 . PMC 4357260 . PMID 25814977 .
- ^ Марино Ф., Косентино М (апрель 2016 г.). «Стипно -склероз: перепрофилирование дофаминергических препаратов для РС - доказательства устанавливаются». Природные обзоры. Неврология . 12 (4): 191–92. doi : 10.1038/nrneurol.2016.33 . PMID 27020558 . S2CID 26319461 .
- ^ Диксон Д.В. (2007). «Невропатология нарушений движения». В Tolosa E, Jankovic JJ (ред.). Болезнь Паркинсона и расстройства движения . Hagerstown, MD: Lippincott Williams & Wilkins. С. 271–83. ISBN 978-0-7817-7881-7 .
- ^ Jump up to: а беременный Tuite PJ, Krawczewski K (апрель 2007 г.). «Паркинсонизм: подход обзора систем к диагностике». Семинары в неврологии . 27 (2): 113–22. doi : 10.1055/s-2007-971174 . PMID 17390256 . S2CID 260319916 .
- ^ Jump up to: а беременный Olsen CM (декабрь 2011 г.). «Естественные награды, нейропластичность и не лекарства» . Нейрофармакология . 61 (7): 1109–22. doi : 10.1016/j.neuropharm.2011.03.010 . PMC 3139704 . PMID 21459101 .
- ^ Ceravolo R, Frosini D, Rossi C, Bonuccelli U (ноябрь 2010 г.). «Спектр зависимости при болезни Паркинсона: от синдрома дизрегуляции дофамина до импульсных нарушений». Журнал неврологии . 257 (Suppl 2): S276–83. doi : 10.1007/s00415-010-5715-0 . PMID 21080189 . S2CID 19277026 .
- ^ Jump up to: а беременный в дюймовый и фон глин Ghodse H (2010). Наркотики Годсе и привыкание: руководство по лечению (4 -е изд.). Издательство Кембриджского университета. С. 87–92. ISBN 978-1-139-48567-8 .
- ^ Сицилиано К.А., Джонс С.Р. (август 2017 г.). «Потенция кокаина в транспортере дофамина отслеживает дискретные мотивационные состояния во время самостоятельного введения кокаина» . Нейропсихофармакология . 42 (9): 1893–1904. doi : 10.1038/npp.2017.24 . PMC 5520781 . PMID 28139678 .
- ^ Heal DJ, Pierce DM (2006). «Метилфенидат и его изомеры: их роль в лечении расстройства гиперактивности с дефицитом внимания с использованием трансдермальной системы доставки». ЦНС лекарства . 20 (9): 713–38. doi : 10.2165/00023210-200620090-00002 . PMID 16953648 . S2CID 39535277 .
- ^ Jump up to: а беременный Freye E (2009). Фармакология и злоупотребление кокаином, амфетаминами, экстазими и связанными с ними дизайнерскими препаратами. Комплексный обзор их способа действия, лечения злоупотреблений и опьянения . Дордрехт: Спрингер. ISBN 978-90-481-2448-0 .
- ^ Jump up to: а беременный Kimko HC, Cross JT, Abernethy DR (декабрь 1999 г.). «Фармакокинетика и клиническая эффективность метилфенидата». Клиническая фармакокинетика . 37 (6): 457–70. doi : 10.2165/00003088-199937060-00002 . PMID 10628897 . S2CID 397390 .
- ^ Mignot EJ (октябрь 2012 г.). «Практическое руководство по терапии синдромов нарколепсии и гиперсомнии» . Нейротерапевтика . 9 (4): 739–52. doi : 10.1007/s13311-012-0150-9 . PMC 3480574 . PMID 23065655 .
- ^ Zimmerman JL (октябрь 2012 г.). «Интоксикация кокаина». Клиники интенсивной терапии . 28 (4): 517–26. doi : 10.1016/j.ccc.2012.07.003 . PMID 22998988 .
- ^ «Квилливант XR - метилфенидат гидрохлорида суспензия, расширенное высвобождение» . Dailemed.nlm.nih.gov . Получено 11 июля 2020 года .
- ^ Лопес Ф.А., Леру -младший (сентябрь 2013 г.). «Стимуляторы длительного действия для лечения дефицита внимания/гиперактивности: акцент на составах с расширенным высвобождением и пролекарством лисдексамфетамина димелат для решения продолжающихся клинических проблем» . Дефицит внимания и расстройства гиперактивности . 5 (3): 249–65. doi : 10.1007/s12402-013-0106-x . PMC 3751218 . PMID 23564273 .
- ^ Nutt DJ, Lingford-Hughes A, Erritzoe D, Stokes PR (май 2015). «Теория зависимости дофамина: 40 лет максимумов и минимумов» (PDF) . Природные обзоры. Нейробиология . 16 (5): 305–12. doi : 10.1038/nrn3939 . PMID 25873042 . S2CID 2055511111 .
- ^ Jump up to: а беременный в Синха р (август 2013 г.). «Клиническая нейробиология жажды лекарств» . Современное мнение о нейробиологии . 23 (4): 649–54. doi : 10.1016/j.conb.2013.05.001 . PMC 3735834 . PMID 23764204 .
- ^ Volkow ND, Baler RD (январь 2014 г.). «Наука с зависимостью: раскрытие нейробиологической сложности» . Нейрофармакология . 76 (Pt B): 235–49. doi : 10.1016/j.neuropharm.2013.05.007 . PMC 3818510 . PMID 23688927 .
- ^ Nestler EJ (декабрь 2012 г.). «Транскрипционные механизмы наркомании» . Клиническая психофармакология и нейробиология . 10 (3): 136–43. doi : 10.9758/cpn.2012.10.3.136 . PMC 3569166 . PMID 23430970 .
- ^ Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Roffman JL, Roffman JW, Zöllei L, Polimeni JR, Fischl B, Liu H, Buckner RL (сентябрь 2011 г.). «Организация коры головного мозга человека, оцененная по внутренней функциональной связности » Журнал нейрофизиологии 106 (3): 1125–6 Doi : 10.1152/ jn.00338.2 3174820PMC 21653723PMID
- ^ Jump up to: а беременный в дюймовый и фон глин Хили Д. (2004). Создание психофармакологии . Гарвардский университет издательство. С. 37–73. ISBN 978-0-674-01599-9 .
- ^ Jump up to: а беременный Брунтон Л. Гудман и Гилман «Фармакологическая основа терапии» (12 -е изд.). МакГроу Хилл. С. 417–55.
- ^ Jump up to: а беременный в дюймовый Howes od, Kapur S (май 2009 г.). «Гипотеза допамина шизофрении: версия III - последний общий путь» . Бюллетень шизофрении . 35 (3): 549–62. doi : 10.1093/schbul/sbp006 . PMC 2669582 . PMID 19325164 .
- ^ Horacek J, Bubenikova-Valesova V, Kopecek M, Palenicek T, Dockery C, Mohr P, Höschl C (2006). «Механизм действия атипичных антипсихотических препаратов и нейробиологии шизофрении». ЦНС лекарства . 20 (5): 389–409. doi : 10.2165/00023210-200620050-00004 . PMID 16696579 . S2CID 18226404 .
- ^ Шталь С.М. (2018). «За пределами дофаминовой гипотезы шизофрении до трех нейронных сетей психоза: дофамин, серотонин и глутамат» (PDF) . CNS Spectr . 23 (3): 187–91. doi : 10.1017/s1092852918001013 . PMID 29954475 . S2CID 49599226 . Архивировано (PDF) из оригинала 29 апреля 2020 года.
- ^ Jump up to: а беременный Malenka RC, Nestler EJ, Hyman SE (2009). «Главы 10 и 13». В Сидоре А, Браун Р.Ю. (ред.). Молекулярная нейрофармакология: основание для клинической нейробиологии (2 -е изд.). Нью-Йорк: McGraw-Hill Medical. С. 266, 318–23. ISBN 978-0-07-148127-4 .
- ^ Wu J, Xiao H, Sun H, Zou L, Zhu LQ (июнь 2012 г.). «Роль дофаминовых рецепторов в СДВГ: систематический метаанализ». Молекулярная нейробиология . 45 (3): 605–20. doi : 10.1007/s12035-012-8278-5 . PMID 22610946 . S2CID 895006 .
- ^ Jump up to: а беременный Берридж CW, Devilbiss DM (июнь 2011 г.). «Психостимуляторы как когнитивные усилители: префронтальная кора, катехоламины и дефицит внимания/гиперактивность» . Биологическая психиатрия . 69 (12): E101–11. doi : 10.1016/j.biopsych.2010.06.023 . PMC 3012746 . PMID 20875636 .
- ^ Спенсер RC, Devilbiss DM, Berridge CW (июнь 2015 г.). «Познавающие познавательные эффекты психостимулянтов включают прямое действие в префронтальной коре» . Биологическая психиатрия . 77 (11): 940–50. doi : 10.1016/j.biopsych.2014.09.013 . PMC 4377121 . PMID 25499957 .
- ^ Ilieva IP, Hook CJ, Farah MJ (июнь 2015 г.). «Рецептурные стимуляторы на здоровый ингибирующий контроль, рабочую память и эпизодическую память: метаанализ» . Журнал когнитивной нейробиологии . 27 (6): 1069–89. doi : 10.1162/jocn_a_00776 . PMID 25591060 . S2CID 15788121 .
- ^ Jump up to: а беременный в Вуд ПБ (май 2008 г.). «Роль центрального дофамина в боли и анальгезии». Экспертный обзор нейротерапевтики . 8 (5): 781–97. doi : 10.1586/14737175.8.5.781 . PMID 18457535 . S2CID 24325199 .
- ^ Jump up to: а беременный в дюймовый Flake Za, Scalley Rd, Bailey AG (март 2004 г.). «Практический отбор противорвозных средств» . Американский семейный врач . 69 (5): 1169–74. PMID 15023018 .
- ^ Connolly BS, Lang AE (2014). «Фармакологическое лечение болезни Паркинсона: обзор». Джама . 311 (16): 1670–83. doi : 10.1001/Jama.2014.3654 . PMID 24756517 .
- ^ Roshchina VV (2010). «Эволюционные соображения нейротрансмиттеров в микробных, растений и клетках животных». В Lyte M, Primrose PE (Eds.). Микробная эндокринология . Нью -Йорк: Спрингер. С. 17–52. ISBN 978-1-4419-5576-0 .
- ^ Jump up to: а беременный Iyer LM, Aravind L, Coon SL, Klein DC, Koonin EV (июль 2004 г.). «Эволюция передачи сигналов клеток у животных: сыграл ли поздний горизонтальный перенос генов из бактерий?». Тенденции в генетике . 20 (7): 292–99. doi : 10.1016/j.tig.2004.05.007 . PMID 15219393 .
- ^ Jump up to: а беременный в дюймовый и Barron AB, Søvik E, Cornish JL (2010). «Роли дофамина и связанных соединений в поведении, обращающемся за вознаграждением, между филами животных» . Границы в поведенческой нейробиологии . 4 : 163. DOI : 10.3389/fnbeh.2010.00163 . PMC 2967375 . PMID 21048897 .
- ^ Liu H, Misshima Y, Fujiwa T, Nagai H, Visual A, Mine Y, T al. (2004). /Xestposts представляют собой алкалоиды и дофамин Арагпонаж . Морские лекарства 2 (4): 154–63. doi : 10.3390/ md2 PMC 3783253 .
- ^ Kass-Simon G, Pierobon P (январь 2007 г.). «Cnidarian Chemical Neurosmission, обновленный обзор». Сравнительная биохимия и физиология. Часть A, молекулярная и интегративная физиология . 146 (1): 9–25. doi : 10.1016/j.cbpa.2006.09.008 . PMID 17101286 .
- ^ Коттрелл Г.А. (январь 1967 г.). «Появление дофамина и норадреналина в нервной ткани некоторых видов беспозвоночных» . Британский журнал фармакологии и химиотерапии . 29 (1): 63–69. doi : 10.1111/j.1476-5381.1967.tb01939.x . PMC 1557178 . PMID 19108240 .
- ^ Kindt KS, Quast KB, Giles AC, DE S, Hendrey D, Nicastro I, Rankin CH, Schafer WR (август 2007 г.). «Дофамин опосредует контекстную зависимую модуляцию сенсорной пластичности у C. elegans» . Нейрон . 55 (4): 662–76. doi : 10.1016/j.neuron.2007.07.023 . PMID 17698017 . S2CID 2092645 .
- ^ Каливас П.В., Стюарт Дж (1 сентября 1991 г.). «Передача дофамина в инициации и экспрессии чувствительности, вызванной лекарственной и стрессом, двигательной активности». Исследование мозга. Обзоры исследований мозга . 16 (3): 223–44. doi : 10.1016/0165-0173 (91) 90007-U . PMID 1665095 . S2CID 10775295 .
- ^ Perry CJ, Barron AB (2013). «Нейронные механизмы вознаграждения у насекомых» (PDF) . Ежегодный обзор энтомологии . 58 (1): 543–62. doi : 10.1146/annurev-ento-120811-153631 . PMID 23020615 . S2CID 19678766 . Архивировано из оригинала (PDF) 5 июня 2020 года.
- ^ Takikawa Y, Kawagoe R, Hikosaka O (октябрь 2004 г.). «Возможная роль дофаминовых нейронов среднего мозга в краткосрочной и долгосрочной адаптации саккад к картированию положения» (PDF) . Журнал нейрофизиологии . 92 (4): 2520–29. doi : 10.1152/jn.00238.2004 . PMID 15163669 . S2CID 12534057 . Архивировано из оригинала (PDF) 2 марта 2019 года.
- ^ Yamagata N, Ichinose T, Aso Y, Plaçais Py, Friedrich AB, Sima RJ, et al. (Январь 2015). «Различные дофаминовые нейроны опосредуют сигналы вознаграждения для краткосрочных и долгосрочных воспоминаний» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (2): 578–83. Bibcode : 2015pnas..112..578y . doi : 10.1073/pnas.1421930112 . PMC 4299218 . PMID 25548178 .
- ^ Jump up to: а беременный Waddell S (июнь 2013 г.). «Передача сигнализации у Drosophila; дофамин делает все это в конце концов» . Современное мнение о нейробиологии . 23 (3): 324–29. doi : 10.1016/j.conb.2013.01.005 . PMC 3887340 . PMID 23391527 .
- ^ Jump up to: а беременный в дюймовый и фон Кулма А., Сзопа Дж. (2007). «Катехоламины являются активными соединениями в растениях». Наука растений . 172 (3): 433–40. Bibcode : 2007plnsc.172..433k . doi : 10.1016/j.plantsci.2006.10.013 .
- ^ Jump up to: а беременный Ингл П.К. (2003). "L-Dopa Planting Plants" (PDF) . Слушание натурального продукта . 2 : 126–33. Архивировано (PDF) из оригинала 2 марта 2014 года . Получено 24 сентября 2015 года .
- ^ Wichers HJ, Visser JF, Huizing HJ, Pras N (1993). «Появление L-DOPA и дофамина в растениях и клеточных культурах мукуны Pruriens и влияние 2, 4-D и NaCl на эти соединения». Растительные клетки, ткани и органная культура . 33 (3): 259–64. doi : 10.1007/bf02319010 . S2CID 44814336 .
- ^ Лонго Р., Кастеллани А., Сберзе П., Тиболла М. (1974). «Распределение L-DOPA и связанных с ними аминокислот в Vicia». Фитохимия . 13 (1): 167–71. Bibcode : 1974pchem..13..167L . doi : 10.1016/s0031-9422 (00) 91287-1 .
- ^ Ван Алстин К.Л., Нельсон А.В., Вайвьян младший, Канцилла Да (июнь 2006 г.). «Дофамин функционирует как защита против Хербвора в умеренной зеленой водоросли Ulvaria obscura» Оэкология 148 (2): 304–1 Bibcode : 2006oecol.148..304V Doi : 10.1007/ s00442-006-0378-3 16489461PMID 5029574S2CID
- ^ Jump up to: а беременный в Саймон Д.Д., Пелес Д., Вакамацу К., Ито С (октябрь 2009 г.). «Современные проблемы в понимании меланогенеза: соединение химии, биологического контроля, морфологии и функций» . Пигментные клеточные и меланому исследования . 22 (5): 563–79. doi : 10.1111/j.1755-148x.2009.00610.x . PMID 19627559 .
- ^ Fedorow H, Tribl F, Halliday G, Gerlach M, Riederer P, Double KL (февраль 2005 г.). «Нейромеланин в нейронах дофамина человека: сравнение с периферическими меланинами и отношение к болезни Паркинсона». Прогресс в нейробиологии . 75 (2): 109–24. doi : 10.1016/j.pneurobio.2005.02.001 . PMID 15784302 . S2CID 503902 .
- ^ Эндрюс Р.С., Придхэм Дж.Б. (1967). «Меланины из допа, содержащих растения». Фитохимия . 6 (1): 13–18. Bibcode : 1967pchem ... 6 ... 13a . doi : 10.1016/0031-9422 (67) 85002-7 .
- ^ Beldade P, Brakefield PM (июнь 2002 г.). «Генетика и эво-дево из узоров крыла бабочки». Природные обзоры. Генетика . 3 (6): 442–52. doi : 10.1038/nrg818 . PMID 12042771 . S2CID 17417235 .
- ^ Фан С. (2008). «История дофамина и леводопа при лечении болезни Паркинсона». Движение расстройства . 23 (Suppl 3): S497–508. doi : 10.1002/mds.22028 . PMID 18781671 . S2CID 45572523 .
- ^ Бенес FM (январь 2001 г.). «Карлссон и открытие дофамина». Тенденции в фармакологических науках . 22 (1): 46–47. doi : 10.1016/s0165-6147 (00) 01607-2 . PMID 11165672 .
- ^ Barondes SH (2003). Лучше, чем прозак . Нью -Йорк: издательство Оксфордского университета. С. 21–22, 39–40 . ISBN 978-0-19-515130-5 .
- ^ Lee H, Dellatore SM, Miller WM, Messersmith PB (октябрь 2007 г.). «Вдохновленный мидий химия поверхности для многофункциональных покрытий» . Наука . 318 (5849): 426–30. Bibcode : 2007sci ... 318..426L . doi : 10.1126/science.1147241 . PMC 2601629 . PMID 17947576 .
- ^ Jump up to: а беременный Dreyer DR, Miller DJ, Freeman BD, Paul DR, Bielawski CW (2013). «Перспективы на поли (дофамин)». Химическая наука . 4 (10): 3796. DOI : 10.1039/C3SC51501J .
- ^ Jump up to: а беременный в дюймовый и Lynge Me, Van der Westen R, Postma A, Städler B (декабрь 2011 г.). «Полидопамин-полимерное покрытие, вдохновленное природой для биомедицинской науки» (PDF) . Наноразмерный . 3 (12): 4916–28. Bibcode : 2011nanos ... 3.4916L . doi : 10.1039/c1nr10969c . PMID 22024699 . Архивировано с оригинала 7 марта 2014 года.
- ^ Fichman G, Schneider JP (март 2021 г.). «Самополимеризация дофамина как простой и мощный инструмент для модуляции вязкоупругих механических свойств гелей на основе пептидов» . Молекулы . 26 (5): 1363. doi : 10.3390/molecules26051363 . PMC 7961423 . PMID 33806346 .
Дальнейшее чтение (самое последнее первое)
[ редактировать ]- Szalavitz M (13 сентября 2024 г.). «« Дофаминовый быстрый »не спасет вас от зависимости» . New York Times .
- Смит Д.Г. (30 июня 2023 г.). «У нас проблема дофамина» . New York Times .
- Мец С (29 сентября 2021 г.). Бесконечная охота на дофамин в виртуальной реальности в виртуальной реальности . New York Times .
- Лембке А (2021). Дофаминовая нация . Заголовок. ISBN 9781472294128 .
Внешние ссылки
[ редактировать ]- Биология дефицита внимания дефицит гиперактивности
- Дофамин
- Сердечные стимуляторы
- Катехоламины
- Дофаминовые агонисты
- Счастливые гормоны
- Гормоны гипоталамуса
- Гормоны оси гипоталамуса-гипофизационного пролактина
- Эндокринная система человека
- Инотропные агенты
- Моноаминергические нейротоксины
- Мотивация
- Нейротрансмиттеры
- Норэпинефрино-дофаминовые агенты
- Периферически селективные лекарства
- Фенотиламины
- Агонисты таар1