Серотонин
 | |
| Клинические данные | |
|---|---|
| Другие имена | 5-HT, 5-гидрокситриптамина, энтермина, тромбоцитина, 3- (β-аминоэтил) -5-гидроксильный раствор, тромботонин |
| Физиологические данные | |
| Исходные ткани | ядра рафе , энтерохромаффиновые клетки |
| Целевые ткани | по всей системе |
| Receptors | 5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6, 5-HT7 |
| Agonists | Indirectly: SSRIs, MAOIs |
| Precursor | 5-HTP |
| Biosynthesis | Aromatic L-amino acid decarboxylase |
| Metabolism | MAO |
| Identifiers | |
| CAS Number | |
| PubChem CID | |
| IUPHAR/BPS | |
| ChemSpider | |
| KEGG | |
| PDB ligand | |
| CompTox Dashboard (EPA) | |
| ECHA InfoCard | 100.000.054 |

| |
| Names | |
|---|---|
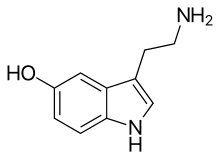
| IUPAC name
5-Hydroxytryptamine
| |
| Preferred IUPAC name
3-(2-Aminoethyl)-1H-indol-5-ol | |
| Other names
5-Hydroxytryptamine, 5-HT, Enteramine; Thrombocytin, 3-(β-Aminoethyl)-5-hydroxyindole, 3-(2-Aminoethyl)indol-5-ol, Thrombotonin
| |
| Identifiers | |
3D model (JSmol)
|
|
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| ECHA InfoCard | 100.000.054 |
| KEGG | |
| MeSH | Serotonin |
PubChem CID
|
|
| UNII | |
CompTox Dashboard (EPA)
|
|
| Properties | |
| C10H12N2O | |
| Molar mass | 176.215 g/mol |
| Appearance | White powder |
| Melting point | 167.7 °C (333.9 °F; 440.8 K) 121–122 °C (ligroin)[3] |
| Boiling point | 416 ± 30 °C (at 760 Torr)[1] |
| slightly soluble | |
| Acidity (pKa) | 10.16 in water at 23.5 °C[2] |
| 2.98 D | |
| Hazards | |
| Lethal dose or concentration (LD, LC): | |
LD50 (median dose)
|
750 mg/kg (subcutaneous, rat),[4] 4500 mg/kg (intraperitoneal, rat),[5] 60 mg/kg (oral, rat) |
| Safety data sheet (SDS) | External MSDS |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
| |
Серотонин ( / ˌ s ɛr ɛ n ɪ n n , ˌ s ɪərə-/ɪɪ [ 6 ] [ 7 ] [ 8 ] или 5-гидрокситриптамина ( 5-HT ) является моноаминовым нейротрансмиттером . Его биологическая функция сложна, затрагивая различные функции, включая настроение , познание , награду , обучение , память и многочисленные физиологические процессы, такие как рвота и вазоконстрикция . [ 9 ]
Серотонин продуцируется в центральной нервной системе (ЦНС), в частности в мозга ствола ядрах кожи , клетки Меркель , легочные нейроэндокринные клетки языка и клетки рецептора вкуса . Приблизительно 90% серотонина, который производит человеческий организм , находится в желудочно -кишечного тракта , энтерохромафиновых клетках где он регулирует движения кишечника. [ 10 ] [ 11 ] [ 12 ] Кроме того, он хранится в тромбоцитах крови и выпускается во время агитации и вазоконстрикции, где он затем действует как агонист в другие тромбоциты. [ 13 ] Около 8% встречаются в тромбоцитах и 1–2% в ЦНС. [ 14 ]
The serotonin is secreted luminally and basolaterally, which leads to increased serotonin uptake by circulating platelets and activation after stimulation, which gives increased stimulation of myenteric neurons and gastrointestinal motility.[15] The remainder is synthesized in serotonergic neurons of the CNS, where it has various functions, including the regulation of mood, appetite, and sleep.[16][unreliable medical source][17][unreliable medical source]
Serotonin secreted from the enterochromaffin cells eventually finds its way out of tissues into the blood. There, it is actively taken up by blood platelets, which store it. When the platelets bind to a clot, they release serotonin, where it can serve as a vasoconstrictor or a vasodilator while regulating hemostasis and blood clotting. In high concentrations, serotonin acts as a vasoconstrictor by contracting endothelial smooth muscle directly or by potentiating the effects of other vasoconstrictors (e.g. angiotensin II and norepinephrine). The vasoconstrictive property is mostly seen in pathologic states affecting the endothelium – such as atherosclerosis or chronic hypertension. In normal physiologic states, vasodilation occurs through the serotonin mediated release of nitric oxide from endothelial cells, and the inhibition of release of norepinephrine from adrenergic nerves.[18] Serotonin is also a growth factor for some types of cells, which may give it a role in wound healing. There are various serotonin receptors.
Biochemically, the indoleamine molecule derives from the amino acid tryptophan. Serotonin is metabolized mainly to 5-hydroxyindoleacetic acid (5-HIAA), chiefly by the liver. Several classes of antidepressants, such as selective serotonin reuptake inhibitors (SSRIs) and serotonin–norepinephrine reuptake inhibitors (SNRIs), interfere with the normal reabsorption of serotonin after it is done with the transmission of the signal, therefore augmenting the neurotransmitter levels in the synapses.
Besides mammals, serotonin is found in all bilateral animals including worms and insects,[19] as well as in fungi and in plants.[20] Serotonin's presence in insect venoms and plant spines serves to cause pain, which is a side-effect of serotonin injection.[21][22] Serotonin is produced by pathogenic amoebae, causing diarrhea in the human gut.[23] Its widespread presence in many seeds and fruits may serve to stimulate the digestive tract into expelling the seeds.[24][failed verification]
Molecular structure
[edit]Biochemically, the indoleamine molecule derives from the amino acid tryptophan, via the (rate-limiting) hydroxylation of the 5 position on the ring (forming the intermediate 5-hydroxytryptophan), and then decarboxylation to produce serotonin.[25] Preferable conformations are defined via ethylamine chain, resulting in six different conformations.[26]
Crystal structure
[edit]Serotonin crystallizes in P212121 chiral space group forming different hydrogen-bonding interactions between serotonin molecules via N-H...O and O-H...N intermolecular bonds.[27] Serotonin also forms several salts, including pharmaceutical formulation of serotonin adipate.[28]
Biological role
[edit]Serotonin is involved in numerous physiological processes,[29] including sleep,[30] thermoregulation, learning and memory, pain, (social) behavior,[31] sexual activity, feeding, motor activity, neural development,[32] and biological rhythms.[33] In less complex animals, such as some invertebrates, serotonin regulates feeding and other processes.[34] In plants serotonin synthesis seems to be associated with stress signals.[20][35] Despite its longstanding prominence in pharmaceutical advertising, the claim that low serotonin levels cause depression is not supported by scientific evidence.[36][37][38]
Cellular effects
[edit]Serotonin primarily acts through its receptors and its effects depend on which cells and tissues express these receptors.[33]
Metabolism involves first oxidation by monoamine oxidase to 5-hydroxyindoleacetaldehyde (5-HIAL).[39][40] The rate-limiting step is hydride transfer from serotonin to the flavin cofactor.[41] There follows oxidation by aldehyde dehydrogenase (ALDH) to 5-hydroxyindoleacetic acid (5-HIAA), the indole acetic-acid derivative. The latter is then excreted by the kidneys.
Receptors
[edit]The 5-HT receptors, the receptors for serotonin, are located on the cell membrane of nerve cells and other cell types in animals, and mediate the effects of serotonin as the endogenous ligand and of a broad range of pharmaceutical and psychedelic drugs. Except for the 5-HT3 receptor, a ligand-gated ion channel, all other 5-HT receptors are G-protein-coupled receptors (also called seven-transmembrane, or heptahelical receptors) that activate an intracellular second messenger cascade.[42]
Termination
[edit]Serotonergic action is terminated primarily via uptake of 5-HT from the synapse. This is accomplished through the specific monoamine transporter for 5-HT, SERT, on the presynaptic neuron. Various agents can inhibit 5-HT reuptake, including cocaine, dextromethorphan (an antitussive), tricyclic antidepressants and selective serotonin reuptake inhibitors (SSRIs). A 2006 study found that a significant portion of 5-HT's synaptic clearance is due to the selective activity of the plasma membrane monoamine transporter (PMAT) which actively transports the molecule across the membrane and back into the presynaptic cell.[43]
In contrast to the high affinity of SERT, the PMAT has been identified as a low-affinity transporter, with an apparent Km of 114 micromoles/l for serotonin, which is approximately 230 times higher than that of SERT. However, the PMAT, despite its relatively low serotonergic affinity, has a considerably higher transport "capacity" than SERT, "resulting in roughly comparable uptake efficiencies to SERT ... in heterologous expression systems."[43] The study also suggests that the administration of SSRIs such as fluoxetine and sertraline may be associated with an inhibitory effect on PMAT activity when used at higher than normal dosages (IC50 test values used in trials were 3–4 fold higher than typical prescriptive dosage).
Serotonylation
[edit]Serotonin can also signal through a nonreceptor mechanism called serotonylation, in which serotonin modifies proteins.[44] This process underlies serotonin's effects upon platelet-forming cells (thrombocytes) in which it links to the modification of signaling enzymes called GTPases that then trigger the release of vesicle contents by exocytosis.[45] A similar process underlies the pancreatic release of insulin.[44]
The effects of serotonin upon vascular smooth muscle tone – the biological function after which serotonin was originally named – depend upon the serotonylation of proteins involved in the contractile apparatus of muscle cells.[46]
| Receptor | Ki (nM)[47] | Receptor function[Note 1] |
|---|---|---|
| 5-HT1 receptor family signals via Gi/o inhibition of adenylyl cyclase. | ||
| 5-HT1A | 3.17 | Memory[vague] (agonists ↓); learning[vague] (agonists ↓); anxiety (agonists ↓); depression (agonists ↓); positive, negative, and cognitive symptoms of schizophrenia (partial agonists ↓); analgesia (agonists ↑); aggression (agonists ↓); dopamine release in the prefrontal cortex (agonists ↑); serotonin release and synthesis (agonists ↓) |
| 5-HT1B | 4.32 | Vasoconstriction (agonists ↑); aggression (agonists ↓); bone mass (↓). Serotonin autoreceptor. |
| 5-HT1D | 5.03 | Vasoconstriction (agonists ↑) |
| 5-HT1E | 7.53 | |
| 5-HT1F | 10 | |
| 5-HT2 receptor family signals via Gq activation of phospholipase C. | ||
| 5-HT2A | 11.55 | Psychedelia (agonists ↑); depression (agonists & antagonists ↓); anxiety (antagonists ↓); positive and negative symptoms of schizophrenia (antagonists ↓); norepinephrine release from the locus coeruleus (antagonists ↑); glutamate release in the prefrontal cortex (agonists ↑); dopamine in the prefrontal cortex (agonists ↑);[48] urinary bladder contractions (agonists ↑)[49] |
| 5-HT2B | 8.71 | Cardiovascular functioning (agonists increase risk of pulmonary hypertension), empathy (via von Economo neurons[50]) |
| 5-HT2C | 5.02 | Dopamine release into the mesocorticolimbic pathway (agonists ↓); acetylcholine release in the prefrontal cortex (agonists ↑); dopaminergic and noradrenergic activity in the frontal cortex (antagonists ↑);[51] appetite (agonists ↓); antipsychotic effects (agonists ↑); antidepressant effects (agonists & antagonists ↑) |
| Other 5-HT receptors | ||
| 5-HT3 | 593 | Emesis (agonists ↑); anxiolysis (antagonists ↑). |
| 5-HT4 | 125.89 | Movement of food across the GI tract (agonists ↑); memory & learning (agonists ↑); antidepressant effects (agonists ↑). Signalling via Gαs activation of adenylyl cyclase. |
| 5-HT5A | 251.2 | Memory consolidation.[52] Signals via Gi/o inhibition of adenylyl cyclase. |
| 5-HT6 | 98.41 | Cognition (antagonists ↑); antidepressant effects (agonists & antagonists ↑); anxiogenic effects (antagonists ↑[53]). Gs signalling via activating adenylyl cyclase. |
| 5-HT7 | 8.11 | Cognition (antagonists ↑); antidepressant effects (antagonists ↑). Acts by Gs signalling via activating adenylyl cyclase. |
Nervous system
[edit]
The neurons of the raphe nuclei are the principal source of 5-HT release in the brain.[54] There are nine raphe nuclei, designated B1–B9, which contain the majority of serotonin-containing neurons (some scientists chose to group the nuclei raphes lineares into one nucleus), all of which are located along the midline of the brainstem, and centered on the reticular formation.[55][56] Axons from the neurons of the raphe nuclei form a neurotransmitter system reaching almost every part of the central nervous system. Axons of neurons in the lower raphe nuclei terminate in the cerebellum and spinal cord, while the axons of the higher nuclei spread out in the entire brain.
Ultrastructure and function
[edit]The serotonin nuclei may also be divided into two main groups, the rostral and caudal containing three and four nuclei respectively. The rostral group consists of the caudal linear nuclei (B8), the dorsal raphe nuclei (B6 and B7) and the median raphe nuclei (B5, B8 and B9), that project into multiple cortical and subcortical structures. The caudal group consists of the nucleus raphe magnus (B3), raphe obscurus nucleus (B2), raphe pallidus nucleus (B1), and lateral medullary reticular formation, that project into the brainstem.[57]
The serotonergic pathway is involved in sensorimotor function, with pathways projecting both into cortical (Dorsal and Median Raphe Nuclei), subcortical, and spinal areas involved in motor activity. Pharmacological manipulation suggests that serotonergic activity increases with motor activity while firing rates of serotonergic neurons increase with intense visual stimuli. Animal models suggest that kainate signaling negatively regulates serotonin actions in the retina, with possible implications for the control of the visual system.[58] The descending projections form a pathway that inhibits pain called the "descending inhibitory pathway" that may be relevant to a disorder such as fibromyalgia, migraine, and other pain disorders, and the efficacy of antidepressants in them.[59]
Serotonergic projections from the caudal nuclei are involved in regulating mood and emotion, and hypo-[60] or hyper-serotonergic[61] states may be involved in depression and sickness behavior.
Microanatomy
[edit]Serotonin is released into the synapse, or space between neurons, and diffuses over a relatively wide gap (>20 nm) to activate 5-HT receptors located on the dendrites, cell bodies, and presynaptic terminals of adjacent neurons.
When humans smell food, dopamine is released to increase the appetite. But, unlike in worms, serotonin does not increase anticipatory behaviour in humans; instead, the serotonin released while consuming activates 5-HT2C receptors on dopamine-producing cells. This halts their dopamine release, and thereby serotonin decreases appetite. Drugs that block 5-HT2C receptors make the body unable to recognize when it is no longer hungry or otherwise in need of nutrients, and are associated with weight gain,[62] especially in people with a low number of receptors.[63] The expression of 5-HT2C receptors in the hippocampus follows a diurnal rhythm,[64] just as the serotonin release in the ventromedial nucleus, which is characterised by a peak at morning when the motivation to eat is strongest.[65]
In macaques, alpha males have twice the level of serotonin in the brain as subordinate males and females (measured by the concentration of 5-HIAA in the cerebrospinal fluid (CSF)). Dominance status and CSF serotonin levels appear to be positively correlated. When dominant males were removed from such groups, subordinate males begin competing for dominance. Once new dominance hierarchies were established, serotonin levels of the new dominant individuals also increased to double those in subordinate males and females. The reason why serotonin levels are only high in dominant males, but not dominant females has not yet been established.[66]
In humans, levels of 5-HT1A receptor inhibition in the brain show negative correlation with aggression,[67] and a mutation in the gene that codes for the 5-HT2A receptor may double the risk of suicide for those with that genotype.[68] Serotonin in the brain is not usually degraded after use, but is collected by serotonergic neurons by serotonin transporters on their cell surfaces. Studies have revealed nearly 10% of total variance in anxiety-related personality depends on variations in the description of where, when and how many serotonin transporters the neurons should deploy.[69]
Outside the nervous system
[edit]Digestive tract (emetic)
[edit]Serotonin regulates gastrointestinal (GI) function. The gut is surrounded by enterochromaffin cells, which release serotonin in response to food in the lumen. This makes the gut contract around the food. Platelets in the veins draining the gut collect excess serotonin. There are often serotonin abnormalities in gastrointestinal disorders such as constipation and irritable bowel syndrome.[70]
If irritants are present in the food, the enterochromaffin cells release more serotonin to make the gut move faster, i.e., to cause diarrhea, so the gut is emptied of the noxious substance. If serotonin is released in the blood faster than the platelets can absorb it, the level of free serotonin in the blood is increased. This activates 5-HT3 receptors in the chemoreceptor trigger zone that stimulate vomiting.[71] Thus, drugs and toxins stimulate serotonin release from enterochromaffin cells in the gut wall. The enterochromaffin cells not only react to bad food but are also very sensitive to irradiation and cancer chemotherapy. Drugs that block 5HT3 are very effective in controlling the nausea and vomiting produced by cancer treatment, and are considered the gold standard for this purpose.[72]
Lungs
[edit]The lung,[73] including that of reptiles,[74] contains specialized epithelial cells that occur as solitary cells or as clusters called neuroepithelial bodies or bronchial Kulchitsky cells or alternatively K cells.[75] These are enterochromaffin cells that like those in the gut release serotonin.[75] Their function is probably vasoconstriction during hypoxia.[73]
Skin
[edit]Serotonin is also produced by Merkel cells which are part of the somatosensory system.[76]
Bone metabolism
[edit]In mice and humans, alterations in serotonin levels and signalling have been shown to regulate bone mass.[77][78][79][80] Mice that lack brain serotonin have osteopenia, while mice that lack gut serotonin have high bone density. In humans, increased blood serotonin levels have been shown to be a significant negative predictor of low bone density. Serotonin can also be synthesized, albeit at very low levels, in the bone cells. It mediates its actions on bone cells using three different receptors. Through 5-HT1B receptors, it negatively regulates bone mass, while it does so positively through 5-HT2B receptors and 5-HT2C receptors. There is very delicate balance between physiological role of gut serotonin and its pathology. Increase in the extracellular content of serotonin results in a complex relay of signals in the osteoblasts culminating in FoxO1/ Creb and ATF4 dependent transcriptional events.[81] Following the 2008 findings that gut serotonin regulates bone mass, the mechanistic investigations into what regulates serotonin synthesis from the gut in the regulation of bone mass have started. Piezo1 has been shown to sense RNA in the gut and relay this information through serotonin synthesis to the bone by acting as a sensor of single-stranded RNA (ssRNA) governing 5-HT production. Intestinal epithelium-specific deletion of mouse Piezo1 profoundly disturbed gut peristalsis, impeded experimental colitis, and suppressed serum 5-HT levels. Because of systemic 5-HT deficiency, conditional knockout of Piezo1 increased bone formation. Notably, fecal ssRNA was identified as a natural Piezo1 ligand, and ssRNA-stimulated 5-HT synthesis from the gut was evoked in a MyD88/TRIF-independent manner. Colonic infusion of RNase A suppressed gut motility and increased bone mass. These findings suggest gut ssRNA as a master determinant of systemic 5-HT levels, indicating the ssRNA-Piezo1 axis as a potential prophylactic target for treatment of bone and gut disorders. Studies in 2008, 2010 and 2019 have opened the potential for serotonin research to treat bone mass disorders.[82][83]
Organ development
[edit]Since serotonin signals resource availability it is not surprising that it affects organ development. Many human and animal studies have shown that nutrition in early life can influence, in adulthood, such things as body fatness, blood lipids, blood pressure, atherosclerosis, behavior, learning, and longevity.[84][85][86] Rodent experiment shows that neonatal exposure to SSRIs makes persistent changes in the serotonergic transmission of the brain resulting in behavioral changes,[87][88] which are reversed by treatment with antidepressants.[89] By treating normal and knockout mice lacking the serotonin transporter with fluoxetine scientists showed that normal emotional reactions in adulthood, like a short latency to escape foot shocks and inclination to explore new environments were dependent on active serotonin transporters during the neonatal period.[90][91]
Human serotonin can also act as a growth factor directly. Liver damage increases cellular expression of 5-HT2A and 5-HT2B receptors, mediating liver compensatory regrowth (see Liver § Regeneration and transplantation)[92] Serotonin present in the blood then stimulates cellular growth to repair liver damage.[93]
5-HT2B receptors also activate osteocytes, which build up bone[94] However, serotonin also inhibits osteoblasts, through 5-HT1B receptors.[95]
Cardiovascular growth factor
[edit]Serotonin, in addition, evokes endothelial nitric oxide synthase activation and stimulates, through a 5-HT1B receptor-mediated mechanism, the phosphorylation of p44/p42 mitogen-activated protein kinase activation in bovine aortic endothelial cell cultures.[clarification needed][96] In blood, serotonin is collected from plasma by platelets, which store it. It is thus active wherever platelets bind in damaged tissue, as a vasoconstrictor to stop bleeding, and also as a fibrocyte mitotic (growth factor), to aid healing.[97]
Pharmacology
[edit]Several classes of drugs target the serotonin system, including some antidepressants, antipsychotics, anxiolytics, antiemetics, and antimigraine drugs, as well as, the psychedelic drugs and entactogens.
Mechanism of action
[edit]At rest, serotonin is stored within the vesicles of presynaptic neurons. When stimulated by nerve impulses, serotonin is released as a neurotransmitter into the synapse, reversibly binding to the postsynaptic receptor to induce a nerve impulse on the postsynaptic neuron. Serotonin can also bind to auto-receptors on the presynaptic neuron to regulate the synthesis and release of serotonin. Normally serotonin is taken back into the presynaptic neuron to stop its action, then reused or broken down by monoamine oxidase.[98]
Psychedelic drugs
[edit]The serotonergic psychedelic drugs psilocin/psilocybin, DMT, mescaline, psychedelic mushroom and LSD are agonists, primarily at 5-HT2A and 5-HT2C receptors.[99][100][101] The empathogen-entactogen MDMA releases serotonin from synaptic vesicles of neurons.[102]
Antidepressants
[edit]Drugs that alter serotonin levels are used in treating depression, generalized anxiety disorder, and social phobia. Monoamine oxidase inhibitors (MAOIs) prevent the breakdown of monoamine neurotransmitters (including serotonin), and therefore increase concentrations of the neurotransmitter in the brain. MAOI therapy is associated with many adverse drug reactions, and patients are at risk of hypertensive emergency triggered by foods with high tyramine content, and certain drugs. Some drugs inhibit the re-uptake of serotonin, making it stay in the synaptic cleft longer. The tricyclic antidepressants (TCAs) inhibit the reuptake of both serotonin and norepinephrine. The newer selective serotonin reuptake inhibitors (SSRIs) have fewer side-effects and fewer interactions with other drugs.[103]
Certain SSRI medications have been shown to lower serotonin levels below the baseline after chronic use, despite initial increases.[104] The 5-HTTLPR gene codes for the number of serotonin transporters in the brain, with more serotonin transporters causing decreased duration and magnitude of serotonergic signaling.[105] The 5-HTTLPR polymorphism (l/l) causing more serotonin transporters to be formed is also found to be more resilient against depression and anxiety.[106][107]
Serotonin syndrome
[edit]Extremely high levels of serotonin can cause a condition known as serotonin syndrome, with toxic and potentially fatal effects. In practice, such toxic levels are essentially impossible to reach through an overdose of a single antidepressant drug, but require a combination of serotonergic agents, such as an SSRI with a MAOI, which may occur in therapeutic doses.[108][109] The intensity of the symptoms of serotonin syndrome vary over a wide spectrum, and the milder forms are seen even at nontoxic levels.[110] It is estimated that 14% of patients experiencing serotonin syndrome overdose on SSRIs; meanwhile the fatality rate is between 2% and 12%.[108][111][112]
Antiemetics
[edit]Some 5-HT3 antagonists, such as ondansetron, granisetron, and tropisetron, are important antiemetic agents. They are particularly important in treating the nausea and vomiting that occur during anticancer chemotherapy using cytotoxic drugs. Another application is in the treatment of postoperative nausea and vomiting.
Other
[edit]Some serotonergic agonist drugs cause fibrosis anywhere in the body, particularly the syndrome of retroperitoneal fibrosis, as well as cardiac valve fibrosis.[113]
In the past, three groups of serotonergic drugs have been epidemiologically linked with these syndromes. These are the serotonergic vasoconstrictive antimigraine drugs (ergotamine and methysergide),[113] the serotonergic appetite suppressant drugs (fenfluramine, chlorphentermine, and aminorex), and certain anti-Parkinsonian dopaminergic agonists, which also stimulate serotonergic 5-HT2B receptors. These include pergolide and cabergoline, but not the more dopamine-specific lisuride.[114]
As with fenfluramine, some of these drugs have been withdrawn from the market after groups taking them showed a statistical increase of one or more of the side effects described. An example is pergolide. The drug was declining in use since it was reported in 2003 to be associated with cardiac fibrosis.[115]
Two independent studies published in The New England Journal of Medicine in January 2007 implicated pergolide, along with cabergoline, in causing valvular heart disease.[116][117] As a result of this, the FDA removed pergolide from the United States market in March 2007.[118] (Since cabergoline is not approved in the United States for Parkinson's Disease, but for hyperprolactinemia, the drug remains on the market. Treatment for hyperprolactinemia requires lower doses than that for Parkinson's Disease, diminishing the risk of valvular heart disease).[119]
Methyl-tryptamines and hallucinogens
[edit]Several plants contain serotonin together with a family of related tryptamines that are methylated at the amino (NH2) and (OH) groups, are N-oxides, or miss the OH group. These compounds do reach the brain, although some portion of them are metabolized by monoamine oxidase enzymes (mainly MAO-A) in the liver. Examples are plants from the genus Anadenanthera that are used in the hallucinogenic yopo snuff. These compounds are widely present in the leaves of many plants, and may serve as deterrents for animal ingestion. Serotonin occurs in several mushrooms of the genus Panaeolus.[120]
Comparative biology and evolution
[edit]Unicellular organisms
[edit]Serotonin is used by a variety of single-cell organisms for various purposes. SSRIs have been found to be toxic to algae.[121] The gastrointestinal parasite Entamoeba histolytica secretes serotonin, causing a sustained secretory diarrhea in some people.[23][122] Patients infected with E. histolytica have been found to have highly elevated serum serotonin levels, which returned to normal following resolution of the infection.[123] E. histolytica also responds to the presence of serotonin by becoming more virulent.[124] This means serotonin secretion not only serves to increase the spread of entamoebas by giving the host diarrhea but also serves to coordinate their behaviour according to their population density, a phenomenon known as quorum sensing. Outside the gut of a host, there is nothing that the entamoebas provoke to release serotonin, hence the serotonin concentration is very low. Low serotonin signals to the entamoebas they are outside a host and they become less virulent to conserve energy. When they enter a new host, they multiply in the gut, and become more virulent as the enterochromaffine cells get provoked by them and the serotonin concentration increases.
Edible plants and mushrooms
[edit]In drying seeds, serotonin production is a way to get rid of the buildup of poisonous ammonia. The ammonia is collected and placed in the indole part of L-tryptophan, which is then decarboxylated by tryptophan decarboxylase to give tryptamine, which is then hydroxylated by a cytochrome P450 monooxygenase, yielding serotonin.[125]
However, since serotonin is a major gastrointestinal tract modulator, it may be produced in the fruits of plants as a way of speeding the passage of seeds through the digestive tract, in the same way as many well-known seed and fruit associated laxatives. Serotonin is found in mushrooms, fruits, and vegetables. The highest values of 25–400 mg/kg have been found in nuts of the walnut (Juglans) and hickory (Carya) genera. Serotonin concentrations of 3–30 mg/kg have been found in plantains, pineapples, banana, kiwifruit, plums, and tomatoes. Moderate levels from 0.1–3 mg/kg have been found in a wide range of tested vegetables.[24][20]
Serotonin is one compound of the poison contained in stinging nettles (Urtica dioica), where it causes pain on injection in the same manner as its presence in insect venoms.[22] It is also naturally found in Paramuricea clavata, or the Red Sea Fan.[126]
Serotonin and tryptophan have been found in chocolate with varying cocoa contents. The highest serotonin content (2.93 μg/g) was found in chocolate with 85% cocoa, and the highest tryptophan content (13.27–13.34 μg/g) was found in 70–85% cocoa. The intermediate in the synthesis from tryptophan to serotonin, 5-hydroxytryptophan, was not found.[127]
Root development in Arabidopsis thaliana is stimulated and modulated by serotonin – in various ways at various concentrations.[128]
Serotonin serves as a plant defense chemical against fungi. When infected with Fusarium crown rot (Fusarium pseudograminearum), wheat (Triticum aestivum) greatly increases its production of tryptophan to synthesize new serotonin.[129] The function of this is poorly understood[129] but wheat also produces serotonin when infected by Stagonospora nodorum – in that case to retard spore production.[130] The model cereal Brachypodium distachyon – used as a research substitute for wheat and other production cereals – also produces serotonin, coumaroyl-serotonin, and feruloyl-serotonin in response to F. graminearum. This produces a slight antimicrobial effect. B. distachyon produces more serotonin (and conjugates) in response to deoxynivalenol (DON)-producing F. graminearum than non-DON-producing.[131] Solanum lycopersicum produces many AA conjugates – including several of serotonin – in its leaves, stems, and roots in response to Ralstonia solanacearum infection.[132]
Invertebrates
[edit]Serotonin functions as a neurotransmitter in the nervous systems of most animals.
Nematodes
[edit]For example, in the roundworm Caenorhabditis elegans, which feeds on bacteria, serotonin is released as a signal in response to positive events, such as finding a new source of food or in male animals finding a female with which to mate.[133] When a well-fed worm feels bacteria on its cuticle, dopamine is released, which slows it down; if it is starved, serotonin also is released, which slows the animal down further. This mechanism increases the amount of time animals spend in the presence of food.[134] The released serotonin activates the muscles used for feeding, while octopamine suppresses them.[135][136] Serotonin diffuses to serotonin-sensitive neurons, which control the animal's perception of nutrient availability.
Decapods
[edit]If lobsters are injected with serotonin, they behave like dominant individuals whereas octopamine causes subordinate behavior.[31] A crayfish that is frightened may flip its tail to flee, and the effect of serotonin on this behavior depends largely on the animal's social status. Serotonin inhibits the fleeing reaction in subordinates, but enhances it in socially dominant or isolated individuals. The reason for this is social experience alters the proportion between serotonin receptors (5-HT receptors) that have opposing effects on the fight-or-flight response.[clarification needed] The effect of 5-HT1 receptors predominates in subordinate animals, while 5-HT2 receptors predominates in dominants.[137]
In venoms
[edit]Serotonin is a common component of invertebrate venoms, salivary glands, nervous tissues, and various other tissues, across molluscs, insects, crustaceans, scorpions, various kinds of worms, and jellyfish.[22] Adult Rhodnius prolixus – hematophagous on vertebrates – secrete lipocalins into the wound during feeding. In 2003 these lipocalins were demonstrated to sequester serotonin to prevent vasoconstriction (and possibly coagulation) in the host.[138]
Insects
[edit]Serotonin is evolutionarily conserved and appears across the animal kingdom. It is seen in insect processes in roles similar to in the human central nervous system, such as memory, appetite, sleep, and behavior.[139][19] Some circuits in mushroom bodies are serotonergic.[140] (See specific Drosophila example below, §Dipterans.)
Acrididae
[edit]Locust swarming is initiated but not maintained by serotonin,[141] with release being triggered by tactile contact between individuals.[142] This transforms social preference from aversion to a gregarious state that enables coherent groups.[143][142][141] Learning in flies and honeybees is affected by the presence of serotonin.[144][145]
Role in insecticides
[edit]Insect 5-HT receptors have similar sequences to the vertebrate versions, but pharmacological differences have been seen. Invertebrate drug response has been far less characterized than mammalian pharmacology and the potential for species selective insecticides has been discussed.[146]
Hymenopterans
[edit]Wasps and hornets have serotonin in their venom,[147] which causes pain and inflammation[21][22] as do scorpions.[148][22] Pheidole dentata takes on more and more tasks in the colony as it gets older, which requires it to respond to more and more olfactory cues in the course of performing them. This olfactory response broadening was demonstrated to go along with increased serotonin and dopamine, but not octopamine in 2006.[149]
Dipterans
[edit]If flies are fed serotonin, they are more aggressive; flies depleted of serotonin still exhibit aggression, but they do so much less frequently.[150] In their crops it plays a vital role in digestive motility produced by contraction. Serotonin that acts on the crop is exogenous to the crop itself and 2012 research suggested that it probably originated in the serotonin neural plexus in the thoracic-abdominal synganglion.[151] In 2011 a Drosophila serotonergic mushroom body was found to work in concert with Amnesiac to form memories.[140] In 2007 serotonin was found to promote aggression in Diptera, which was counteracted by neuropeptide F – a surprising find given that they both promote courtship, which is usually similar to aggression in most respects.[140]
Vertebrates
[edit]Serotonin, also referred to as 5-hydroxytryptamine (5-HT), is a neurotransmitter most known for its involvement in mood disorders in humans. It is also a widely present neuromodulator among vertebrates and invertebrates.[152] Serotonin has been found having associations with many physiological systems such as cardiovascular, thermoregulation, and behavioral functions, including: circadian rhythm, appetite, aggressive and sexual behavior, sensorimotor reactivity and learning, and pain sensitivity.[153] Serotonin's function in neurological systems along with specific behaviors among vertebrates found to be strongly associated with serotonin will be further discussed. Two relevant case studies are also mentioned regarding serotonin development involving teleost fish and mice.
In mammals, 5-HT is highly concentrated in the substantia nigra, ventral tegmental area and raphe nuclei. Lesser concentrated areas include other brain regions and the spinal cord.[152] 5-HT neurons are also shown to be highly branched, indicating that they are structurally prominent for influencing multiple areas of the CNS at the same time, although this trend is exclusive solely to mammals.[153]
5-HT system in vertebrates
[edit]Vertebrates are multicellular organisms in the phylum Chordata that possess a backbone and a nervous system. This includes mammals, fish, reptiles, birds, etc. In humans, the nervous system is composed of the central and peripheral nervous system, with little known about the specific mechanisms of neurotransmitters in most other vertebrates. However, it is known that while serotonin is involved in stress and behavioral responses, it is also important in cognitive functions.[152] Brain organization in most vertebrates includes 5-HT cells in the hindbrain.[152] In addition to this, 5-HT is often found in other sections of the brain in non-placental vertebrates, including the basal forebrain and pretectum.[154] Since location of serotonin receptors contribute to behavioral responses, this suggests serotonin is part of specific pathways in non-placental vertebrates that are not present in amniotic organisms.[155] Teleost fish and mice are organisms most often used to study the connection between serotonin and vertebrate behavior. Both organisms show similarities in the effect of serotonin on behavior, but differ in the mechanism in which the responses occur.
Dogs / canine species
[edit]There are few studies of serotonin in dogs. One study reported serotonin values were higher at dawn than at dusk.[156] In another study, serum 5-HT levels did not seem to be associated with dogs' behavioural response to a stressful situation.[157] Urinary serotonin/creatinine ratio in bitches tended to be higher 4 weeks after surgery. In addition, serotonin was positively correlated with both cortisol and progesterone but not with testosterone after ovariohysterectomy.[158]
Teleost fish
[edit]Like non-placental vertebrates, teleost fish also possess 5-HT cells in other sections of the brain, including the basal forebrain.[154] Danio rerio (zebra fish) are a species of teleost fish often used for studying serotonin within the brain. Despite much being unknown about serotonergic systems in vertebrates, the importance in moderating stress and social interaction is known.[159] It is hypothesized that AVT and CRF cooperate with serotonin in the hypothalamic-pituitary-interrenal axis.[154] These neuropeptides influence the plasticity of the teleost, affecting its ability to change and respond to its environment. Subordinate fish in social settings show a drastic increase in 5-HT concentrations.[159] High levels of 5-HT long term influence the inhibition of aggression in subordinate fish.[159]
Mice
[edit]Researchers at the Department of Pharmacology and Medical Chemistry used serotonergic drugs on male mice to study the effects of selected drugs on their behavior.[160] Mice in isolation exhibit increased levels of agonistic behavior towards one another. Results found that serotonergic drugs reduce aggression in isolated mice while simultaneously increasing social interaction.[160] Each of the treatments use a different mechanism for targeting aggression, but ultimately all have the same outcome. While the study shows that serotonergic drugs successfully target serotonin receptors, it does not show specifics of the mechanisms that affect behavior, as all types of drugs tended to reduce aggression in isolated male mice.[160] Aggressive mice kept out of isolation may respond differently to changes in serotonin reuptake.
Behavior
[edit]Like in humans, serotonin is extremely involved in regulating behavior in most other vertebrates. This includes not only response and social behaviors, but also influencing mood. Defects in serotonin pathways can lead to intense variations in mood, as well as symptoms of mood disorders, which can be present in more than just humans.
Social interaction
[edit]Одним из наиболее исследованных аспектов социального взаимодействия, в котором участвует серотонин, является агрессия. Агрессия регулируется системой 5-HT, поскольку уровни серотонина могут индуцировать или ингибировать агрессивное поведение, как видно у мышей (см. Раздел о мышах) и крабов. [160] While this is widely accepted, it is unknown if serotonin interacts directly or indirectly with parts of the brain influencing aggression and other behaviors.[152] Studies of serotonin levels show that they drastically increase and decrease during social interactions, and they generally correlate with inhibiting or inciting aggressive behavior.[161] The exact mechanism of serotonin influencing social behaviors is unknown, as pathways in the 5-HT system in various vertebrates can differ greatly.[152]
Ответ на стимулы
[ редактировать ]Серотонин важен в путях ответа окружающей среды, наряду с другими нейротрансмиттерами . [ 162 ] В частности, было обнаружено, что он участвует в слуховой обработке в социальных условиях, поскольку первичные сенсорные системы связаны с социальными взаимодействиями. [ 163 ] Серотонин встречается в структуре IC среднего мозга, которая обрабатывает специфические и неспецифические социальные взаимодействия и вокализации. [ 163 ] Он также получает акустические прогнозы, которые передают сигналы в области слуховой обработки. [ 163 ] Исследования предположили, что серотонин формирует слуховую информацию, полученную IC и, следовательно, влияет на ответы на слуховые стимулы. [ 163 ] Это может повлиять на то, как организм реагирует на звуки хищных или других эффектных видов в их окружающей среде, поскольку поглощение серотонина может влиять на агрессию или социальное взаимодействие.
Настроение
[ редактировать ]Мы можем описать настроение не как специфическое для эмоционального статуса, но как связано с относительно длительным эмоциональным состоянием. Ассоциация серотонина с настроением наиболее известна различными формами депрессии и биполярными расстройствами у людей. [ 153 ] Расстройства, вызванные серотонинергической активностью, потенциально способствуют многим симптомам большой депрессии, такими как общее настроение, активность, суицидальные мысли и сексуальная и когнитивная дисфункция . Селективные ингибиторы обратного захвата серотонина (SSRI) представляют собой класс лекарств, которые, как показали, являются эффективным лечением при крупном депрессивном расстройстве и являются наиболее назначенным классом антидепрессантов. Функция SSRI заключается в блокировании обратного захвата серотонина, что делает больше серотонина доступным для поглощения приемного нейрона. Животные были изучены десятилетиями, чтобы понять депрессивное поведение среди видов. Одно из самых знакомых исследований, тест принудительного плавания (FST), был проведен для измерения потенциальной активности антидепрессантов. [ 153 ] Крыс помещали в неизбежный контейнер с водой, в этот момент время, затрачиваемое на неподвижное, и количество активных поведений (таких как брызги или лазание) сравнивались до и после введения панели антидепрессантов. Было показано, что антидепрессанты, которые избирательно ингибируют обратный захват NE, снижают неподвижность и избирательно увеличивают скалолазание, не затрагивая плавание. Тем не менее, результаты СИОЗС также показывают снижение неподвижности, но увеличение плавания, не влияя на скалолазание. Это исследование продемонстрировало важность поведенческих тестов для антидепрессантов, поскольку они могут обнаруживать лекарства, влияя на поведение ядра, а также поведенческие компоненты видов. [ 153 ]
Рост и размножение
[ редактировать ]В нематоде C. elegans искусственное истощение серотонина или увеличение поведения осьутопаминовых сигналов, типичное для низкопродорной среды: C. elegans становится более активным, а спаривание и яйцо подавляется, в то время как противоположное происходит, если серотонин увеличен или октопамин уменьшается у этого животного. [ 34 ] Серотонин необходим для нормального поведения в браке с нематодой, [ 164 ] и склонность оставить еду для поиска партнера. [ 165 ] Серотонинергическая передача сигналов, используемая для адаптации поведения червя к быстрым изменениям в окружающей среде, влияет на инсулиноподобную передачу сигналов и бета -сигнальную путя TGF ,, как это [ 166 ] которые контролируют долгосрочную адапцию.
В фруктовой мух инсулин регулирует сахар в крови , так и действует как фактор роста . Таким образом, в фруктовой мух серотонинергические нейроны регулируют размер тела взрослого, влияя на секрецию инсулина. [ 167 ] [ 168 ] Серотонин также был идентифицирован как триггер для поведения роя в саранчке. [ 143 ] У людей, хотя инсулин регулирует сахар в крови, а IGF регулирует рост, серотонин контролирует высвобождение обоих гормонов, модулируя высвобождение инсулина из бета -клеток в поджелудочной железе посредством серотонилирования сигнальных белков GTPase. [ 44 ] Воздействие СИОЗС во время беременности снижает рост плода. [ 169 ]
Генетически измененные черви C. elegans , у которых отсутствует серотонин, имеют повышенную репродуктивную продолжительность жизни, могут стать ожирением, а иногда и с арестованным развитием в состоянии бездействия . [ 170 ] [ 171 ]
Старение и возрастные фенотипы
[ редактировать ]Известно, что серотонин регулирует старение, обучение и память. Первое доказательство исходит из изучения долголетия у C. elegans . [ 166 ] На ранней стадии старения [ нечеткий ] , уровень серотонина увеличивается, что изменяет локомоторивое поведение и ассоциативную память. [ 172 ] Эффект восстанавливается мутациями и препаратами (включая миансерин и метиотепин ), которые ингибируют серотониновые рецепторы . Наблюдение не противоречит представлению о том, что уровень серотонина падает у млекопитающих и людей, что обычно наблюдается в позднем, но не рано [ нечеткий ] Фаза старения.
Биохимические механизмы
[ редактировать ]Биосинтез
[ редактировать ]
У животных и людей серотонин синтезируется из аминокислоты L - триптофана с помощью короткого метаболического пути, состоящего из двух ферментов , триптофана гидроксилазы (TPH) и ароматической аминокислотной декарбоксилазы (DDC) и коэнзисного пиридоксаль -фосфата . TPH-опосредованная реакция является ограничивающей скоростью этап на пути.
Было показано, что TPH существует в двух формах: TPH1 , обнаруженная в нескольких тканях , и TPH2 , который является нейрон-специфической изоформой . [ 173 ]
Серотонин может быть синтезирован из триптофана в лаборатории с использованием Aspergillus Niger и Psilocybe Coprophila в качестве катализаторов. Первая фаза до 5-гидрокситриптофана потребует, чтобы триптофан сидел в этаноле и воде в течение 7 дней, а затем смешивание в достаточном количестве HC (или другой кислоте) довести pH до 3, а затем добавление NaOH, чтобы сделать pH 13 в течение 1 часа. Полем Aspergillus Niger станет катализатором для этого первого этапа. Вторая фаза для синтеза самого триптофана от 5-гидрокситриптофанского промежуточного соединения потребует добавления этанола и воды и позволить сидеть в течение 30 дней на этот раз. Следующие два шага будут такими же, как и первая фаза: добавление HCl, чтобы сделать pH = 3, а затем добавить NaOH, чтобы сделать pH очень простым в 13 в течение 1 часа. В этой фазе используется Psilocybe Coprophila в качестве катализатора реакции. [ 174 ]

Серотонин, взятый перорально, не проходит в серотонинергические пути центральной нервной системы, потому что он не пересекает барьер крови -мрачного . [ 9 ] Тем не менее, триптофан и его метаболит 5-гидрокситриптофан (5-HTP), от которого синтезируется серотонин, пересекают барьер крови-мозга. Эти агенты доступны в качестве пищевых добавок и в различных продуктах питания и могут быть эффективными серотонинергическими агентами.
Одним из продуктов серотонина является 5-гидроксиндолетическая кислота (5-HIAA), которая выделяется в моче . Серотонин и 5-HIAA иногда вырабатываются в избыточных количествах определенными опухолями или раком , и уровни этих веществ могут измеряться в моче для проверки этих опухолей.
Аналитическая химия
[ редактировать ]Оксид индийного олова рекомендуется для электродного материала при электрохимическом исследовании концентраций, полученных, обнаруженных или потребляемых микробами . [ 175 ] Метод масс -спектрометрии была разработана в 1994 году для измерения молекулярной массы как природных, так и синтетических серотонинов. [ 176 ]
История и этимология
[ редактировать ]Более века было известно, что вазоконстрикторный материал появляется в сыворотке, когда кровь разрешалась сдвигаться. [ 177 ] В 1935 году итальянский Vittorio Erspamer показал экстракт из энтерохромаффиновых клеток, которые сокращались кишечник. Некоторые полагали, что в нем содержался адреналин , но два года спустя Эрспамер смог показать, что это была ранее неизвестная амин , которую он назвал «Энтемин». [ 178 ] [ 179 ] В 1948 году Морис М. Доклад , Арда Грин и Ирвин Пейдж обнаружили клиники Кливленда вазоконстрикторное вещество в крови сыворотки , и, поскольку это был сывороточный агент, влияющий на сосудистый тонус, они назвали его серотонином. [ 180 ]
В 1952 году было показано, что энтехлин является тем же веществом, что и серотонин, и, поскольку широкий спектр физиологических ролей был выяснен, аббревиатура 5-HT правильного химического названия 5-гидроксиптамина стала предпочтительным названием в фармакологическом поле. [ 181 ] Синонимы серотонина включают в себя: 5-гидрокситриптамин, энтермина, вещества DS и 3- (β-аминоэтил) -5-гидроксиндол. [ 182 ] В 1953 году Бетти Тварог и Пейдж обнаружили серотонин в центральной нервной системе. [ 183 ] Пейдж рассмотрел работу Эрспамера над осьминожным Vulgaris , Discoglossus Pictus , Hexaplex Trunculus , Bolinus brandaris , Sepia , Mytilus и Ostrea как действительный и фундаментальный для понимания этого вновь идентифицированного вещества, но считал его более ранние результаты в различных моделях - особенно от крови крысы - к крови крысы - к крови крысы - к крови крысы - к крови крысы - к крови крысы - к крови крысы - к крови крысы - к крови крысы - до крови Будьте слишком смущены присутствием других биологически активных химических веществ, в том числе некоторых других вазоактивных веществ. [ 184 ]
Примечания
[ редактировать ]- ^ Ссылки на функции этих рецепторов доступны на страницах Википедии для конкретного рецептора.
Ссылки
[ редактировать ]- ^ Рассчитано с использованием программного обеспечения Advanced Chemistry Development (ACD/LABS) v11.02 (© 1994–2011 ACD/LABS)
- ^ Mazák K, Dóczy V, Kökösi J, Nosál B (апрель 2009 г.). «Протон-специальная и микроспекция серотонина и 5-гидрокситриптофан». Химия и биоразнообразие . 6 (4): 578–590. Doi : 10.1002/cbdv.200800087 . PMID 19353542 . S2CID 20543931 .
- ^ Shift S (1958). «[Adrice Hii. II. В Ньюшротонине синтезирует]». Farmac; Научное издание (в Италии). 13 (1): 75–79. PMID 1354273 .
- ^ Erspamer V (1952). «Предварительные поиски об индолалхиламинине и на фейлалхиламинине из экстрактов кожи амфибий». Научные исследования . 22 : 694–702.
- ^ Таммисто Т. (1967). «Повышенная токсичность 5-гидрокситриптамина этанолом у крыс и мышей». Annales Medicinae Experimentalis et Biologiae Fenniae . 46 (3): 382–384. PMID 5734241 .
- ^ Джонс Д. (2003) [1917]. Roach P, Hartmann J, Setter J (Eds.). Английский произношенный словарь . Кембридж: издательство Кембриджского университета. ISBN 978-3-12-539683-8 .
- ^ "Серотонин" . Dictionary.com unabridged (онлайн). н.д.
- ^ "Серотонин" . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ Jump up to: а беременный Молодой SN (ноябрь 2007 г.). «Как увеличить серотонин в человеческом мозге без наркотиков» . Журнал психиатрии и нейробиологии . 32 (6): 394–399. PMC 2077351 . PMID 18043762 .
- ^ «Микробы помогают производить серотонин в кишечнике» . Калифорнийский технологический институт . 9 апреля 2015 года . Получено 3 июня 2022 года .
- ^ Король MW. "Серотонин" . Страница медицинской биохимии . Медицинский факультет Индианы . Получено 1 декабря 2009 года .
- ^ Бергер М., Грей Дж., Рот Б.Л. (2009). «Расширенная биология серотонина» . Ежегодный обзор медицины . 60 : 355–366. doi : 10.1146/annurev.med.60.042307.110802 . PMC 5864293 . PMID 19630576 .
- ^ Schlienger RG, Meier CR (2003). «Влияние селективных ингибиторов обратного захвата серотонина на активацию тромбоцитов: могут ли они предотвратить острый инфаркт миокарда?». Американский журнал сердечно -сосудистых лекарств . 3 (3): 149–162. doi : 10.2165/00129784-200303030-00001 . PMID 14727927 . S2CID 23986530 .
- ^ Kling A (2013). 5-HT2A: рецептор серотонина с возможной ролью в заболеваниях суставов (PDF) (тезис). Umeå Universitet. ISBN 978-91-7459-549-9 .
- ^ Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, et al. (Апрель 2015). «Бактерии коренных народов из кишечной микробиоты регулируют биосинтез серотонина хозяина» . Клетка . 161 (2): 264–276. doi : 10.1016/j.cell.2015.02.047 . PMC 4393509 . PMID 25860609 .
- ^ Sangare A, Dubourget R, Geoffroy H, Gallopin T, Rancillac A (октябрь 2016 г.). «Серотонин дифференциально модулирует возбуждающие и ингибирующие синаптические входы в предполагаемые нейроны, способствующие сна, вентролатерального преоптического ядра» (PDF) . Нейрофармакология . 109 : 29–40. doi : 10.1016/j.neuropharm.2016.05.015 . PMID 27238836 .
- ^ Rancillac A (ноябрь 2016 г.). «Серотонин и способные сна нейроны» . Oncotarget . 7 (48): 78222–78223. doi : 10.18632/oncotarget.13419 . PMC 5346632 . PMID 27861160 .
- ^ Vanhoutte PM (февраль 1987 г.). «Серотонин и сосудистая стена». Международный журнал кардиологии . 14 (2): 189–203. doi : 10.1016/0167-5273 (87) 90008-8 . PMID 3818135 .
- ^ Jump up to: а беременный Husser A, Rohwedder A, Apostolopoulou AA, Widmann A, Pfitzenmaier JE, Maiolo EM, et al. (2012). Zars T (ред.). «Серотонинергическая центральная нервная система личинки дрозофилы: анатомия и поведенческая функция» . Plos один . 7 (10): E47518. BIBCODE : 2012PLOSO ... 747518H . doi : 10.1371/journal.pone.0047518 . PMC 3474743 . PMID 23082175 .
- ^ Jump up to: а беременный в Рамакришна А., Гиридхар П., Равишанкар Г.А. (июнь 2011 г.). «Фитозеротонин: обзор» . Сигнализация и поведение растения . 6 (6): 800–809. Bibcode : 2011plsib ... 6..800a . doi : 10.4161/psb.6.6.15242 . PMC 3218476 . PMID 21617371 .
- ^ Jump up to: а беременный Chen J, Lariviere WR (октябрь 2010 г.). «Ноцицептивные и анти-ноцицептивные эффекты инъекции и терапии пчелиного яда: обоюдоострый меч» . Прогресс в нейробиологии . 92 (2): 151–183. doi : 10.1016/j.pneurobio.2010.06.006 . PMC 2946189 . PMID 20558236 .
- ^ Jump up to: а беременный в дюймовый и Erspamer V (1966). «Возникновение индоаликиламинов в природе». 5-гидрокситриптамина и связанные с ними индолелизамины . Берлин , Гейдельберг : Спрингер Берлин Гейдельберг . С. 132–181. doi : 10.1007/978-3-642-85467-5_4 . ISBN 978-3-642-85469-9 .
- ^ Jump up to: а беременный McGowan K, Kane A, Asarkof N, Wicks J, Guerina V, Kellum J, et al. (Август 1983). «Entamoeba histolytica вызывает кишечную секрецию: роль серотонина». Наука . 221 (4612): 762–764. Bibcode : 1983sci ... 221..762m . doi : 10.1126/science.6308760 . PMID 6308760 .
- ^ Jump up to: а беременный Фельдман Дж. М., Ли Эм (октябрь 1985 г.). «Содержание серотонина в пищевых продуктах: влияние на экскрецию мочи 5-гидроксиндоделиной кислоты» . Американский журнал клинического питания . 42 (4): 639–643. doi : 10.1093/ajcn/42.4.639 . PMID 2413754 .
- ^ González-Flores D, Velardo B, Garrido M, et al. (2011). «Приглашение японских слив (Prunus salicina Lindl. Cv. Crimson Globe) увеличивает 6-сульфатоксимелатонин мочи и общий уровень антиоксидантной способности у молодых, средних и пожилых людей: питательные и функциональные характеристики их содержания» . Журнал исследований пищи и питания . 50 (4): 229–236.
- ^ Ричков Д.А., Хантер С., Ковальский В.Ю., Ломзов А.А., Пулхам К.Р., Болдирева Е.В. (июль 2016 г.). «На пути к пониманию кристаллизации из решения. Исследования DFT многокомпонентных кристаллов серотонина». Вычислительная и теоретическая химия . 1088 : 52–61. doi : 10.1016/j.comptc.2016.04.027 .
- ^ Naeem M, Chadeayne AR, Golen JA, Manke DR (апрель 2022 г.). «Кристаллическая структура серотонина» . Acta Crystallographica Раздел e . 78 (Pt 4): 365–368. Bibcode : 2022accre..78..365n . doi : 10.1107/s2056989022002559 . PMC 8983975 . PMID 35492269 .
- ^ Rychkov D, Boldyreva EV, Tumanov NA (сентябрь 2013 г.). «Новая структура соли серотонина: сравнение и конформационный анализ всех известных серотониновых комплексов». Acta Crystallographica Раздел c . 69 (Pt 9): 1055–1061. BIBCODE : 2013ACCRC..69.1055R . doi : 10.1107/s0108270113019823 . PMID 24005521 .
- ^ Мохаммад-Задех Л.Ф., Моисей Л., Гвалтни-Брант С.М. (июнь 2008 г.). «Серотонин: обзор» . Журнал ветеринарной фармакологии и терапии . 31 (3): 187–199. doi : 10.1111/j.1365-2885.2008.00944.x . PMID 18471139 .
- ^ Vaseghi S, Arjmandi-Rad S, Eskandari M, Ebrahimnejad M, Kholghi G, Zarrindast MR (февраль 2022 г.). «Модулируя роль серотонинергической передачи сигналов во сне и памяти». Фармакологические отчеты . 74 (1): 1–26. doi : 10.1007/s43440-021-00339-8 . PMID 34743316 .
- ^ Jump up to: а беременный Кравиц EA (сентябрь 1988 г.). «Гормональный контроль над поведением: амины и смещение поведенческого производства у омаров». Наука . 241 (4874): 1775–1781. Bibcode : 1988sci ... 241.1775K . doi : 10.1126/science.2902685 . PMID 2902685 .
- ^ Sinclair-Wilson A, Lawrence A, Ferezou I, Cartonnet H, Mailhes C, Garel S, et al. (Август 2023). «Пластичность таламокортикальных аксонов регулируется уровнями серотонина, модулируемых преждевременными рождением» . Труды Национальной академии наук Соединенных Штатов Америки . 120 (33): E23016444120. Bibcode : 2023pnas..12001644S . doi : 10.1073/pnas.23016444120 . PMC 10438379 . PMID 37549297 .
- ^ Jump up to: а беременный Zifa E, Filion G (сентябрь 1992 г.). «5-гидрокситриптаминовые рецепторы» . Фармакологические обзоры . 44 (3): 401–458. PMID 1359584 .
- ^ Jump up to: а беременный Шринивасан С., Садег Л., Эль -И.К., Кристенсен А.Г., Фаермен Н.Дж., Ашрафи К (июнь 2008 г.). «Серотонин регулирует жир C. elegans и питание через независимые молекулярные механизмы» . Клеточный метаболизм . 7 (6): 533–544. doi : 10.1016/j.cmet.2008.04.012 . PMC 2495008 . PMID 18522834 .
- ^ Рамакришна А., Равишанкар Г.А. (ноябрь 2011 г.). «Влияние сигналов абиотического стресса на вторичные метаболиты у растений» . Сигнализация и поведение растения . 6 (11). Информация : 1720–1731. Bibcode : 2011plsib ... 6.1720a . doi : 10.4161/psb.6.11.17613 . PMC 3329344 . PMID 22041989 .
- ^ Whitaker R, Cosgrove L (2015). Психиатрия под влиянием: институциональная коррупция, социальные травмы и рецепты на реформу . Спрингер. С. 55–56. ISBN 978-1-137-51602-2 .
- ^ Moncrieff J, Cooper RE, Stockmann T, Amendola S, Hengartner MP, Горовиц М.А. (август 2023 г.). «Теория депрессии серотонина: систематический обзор доказательств» . Молекулярная психиатрия . 28 (8). Nature Publishing Group: 3243–3256. doi : 10.1038/s41380-022-01661-0 . PMC 10618090 . PMID 35854107 . S2CID 250646781 .
- ^ GHAEMI N (2022), была ли развенчана гипотеза серотонина? , Получено 2 мая 2023 г.
- ^ Bortolato M, Chen K, Shih JC (2010). «Разложение серотонина: роль Мао». Справочник по поведенческой нейробиологии . Тол. 21. Elsevier. п. 203–218. doi : 10.1016/s1569-7339 (10) 70079-5 . ISBN 978-0-12-374634-4 .
- ^ Matthes S, Mosienko V, Bashammakh S, Alenina N, Bader M (2010). «Триптофан -гидроксилаза как новая мишень для лечения депрессивных расстройств». Фармакология . 85 (2): 95–109. doi : 10.1159/000279322 . PMID 20130443 .
- ^ Prah A, Purg M, Stare J, Vianello R, Mavri J (сентябрь 2020 г.). «Как моноаминоксидаза разлагает серотонин: эмпирическое моделирование валентной связи реактивного шага» . Журнал физической химии б . 124 (38): 8259–8265. doi : 10.1021/acs.jpcb.0c06502 . PMC 7520887 . PMID 32845149 .
- ^ Хэннон Дж., Хойер Д. (декабрь 2008 г.). «Молекулярная биология рецепторов 5-HT». Поведенческое исследование мозга . 195 (1): 198–213. doi : 10.1016/j.bbr.2008.03.020 . PMID 18571247 . S2CID 46043982 .
- ^ Jump up to: а беременный Чжоу М., Энгель К, Ван Дж (январь 2007 г.). «Доказательства значительного вклада недавно выявленного моноаминового транспортера (PMAT) в поглощение серотонина в человеческом мозге» . Биохимическая фармакология . 73 (1): 147–154. doi : 10.1016/j.bcp.2006.09.008 . PMC 1828907 . PMID 17046718 .
- ^ Jump up to: а беременный в Paulmann N, Grohmann M, Voigt JP, Bert B, Vowinckel J, Bader M, et al. (Октябрь 2009 г.). O'Rahilly S (ред.). «Внутриклеточный серотонин модулирует секрецию инсулина из бета-клеток поджелудочной железы белком серотонилированием» . PLOS Биология . 7 (10): E1000229. doi : 10.1371/journal.pbio.1000229 . PMC 2760755 . PMID 19859528 .
- ^ Walther DJ, Peter Ju, Winter S, Höltje M, Paulmann N, Grohmann M, et al. (Декабрь 2003 г.). «Серотонилирование небольших GTPases-это путь передачи сигнала, который запускает высвобождение альфа-гранулы тромбоцитов» . Клетка . 115 (7): 851–862. doi : 10.1016/s0092-8674 (03) 01014-6 . PMID 14697203 . S2CID 16847296 .
- ^ Watts SW, Priestley Jr, Thompson JM (май 2009 г.). «Серотонилирование сосудистых белков, важных для сокращения» . Plos один . 4 (5): E5682. Bibcode : 2009ploso ... 4.5682w . doi : 10.1371/journal.pone.0005682 . PMC 2682564 . PMID 19479059 .
- ^ Рот Бл, Дрискол Дж. (12 января 2011 г.). "PDSP K I Database" . Психоактивная программа скрининга лекарств (PDSP) . Университет Северной Каролины в Чапел -Хилл и Национальный институт психического здоровья США. Архивировано с оригинала 8 ноября 2013 года . Получено 17 декабря 2013 года .
- ^ Bortolozzi A, Díaz-Mataix L, Scorza MC, Celada P, Artigas F (декабрь 2005 г.). «Активация рецепторов 5-HT в префронтальной коре усиливает дофаминергическую активность». Журнал нейрохимии . 95 (6): 1597–1607. Doi : 10.1111/j.1471-4159.2005.03485.x . HDL : 10261/33026 . PMID 16277612 . S2CID 18350703 .
- ^ Моро С., Эдвардс Л., Шахматы-Уильямс Р. (ноябрь 2016 г.). «Усовершенствование рецептора 5-HT 2A сократительной активности свиной уротелия и пластинки Propria» . Международный журнал урологии . 23 (11): 946–951. doi : 10.1111/iju.13172 . PMID 27531585 .
- ^ "Von Ecounly Neuon - Neuronbank" . Neuronbank.org . [ ненадежный медицинский источник? ]
- ^ Миллан М.Дж., Гобер А., Лежен Ф., Декин А., Ньюман-Танкреди А., Пастоу В. и др. (Сентябрь 2003 г.). «Роман мелатонинового агониста агомелатина (S20098) является антагонистом в рецепторах 5-гидрокситриптамин2C, блокада которых усиливает активность лобнокортикальных дофаминергических и адренергических путей». Журнал фармакологии и экспериментальной терапии . 306 (3): 954–964. doi : 10.1124/jpet.103.051797 . PMID 12750432 . S2CID 18753440 .
- ^ Гонсалес Р., Чавес-Паскасио К., Менесес А (сентябрь 2013 г.). «Роль рецепторов 5-HT5A в консолидации памяти». Поведенческое исследование мозга . 252 : 246–251. doi : 10.1016/j.bbr.2013.05.051 . PMID 23735322 . S2CID 140204585 .
- ^ Nautiyal KM, Hen R (2017). «Рецепторы серотонина при депрессии: от А до В» . F1000Research . 6 : 123. doi : 10.12688/f1000research.9736.1 . PMC 5302148 . PMID 28232871 .
- ^ Фрэзер А., Хенслер Дж.Г. (1999). «Понимание нейроанатомической организации серотонинергических клеток в мозге дает представление о функциях этого нейротрансмиттера» . В Siegel GJ , Agranoff, Bernard W, Fisher SK, Albers RW, Uhler MD (ред.). Основная нейрохимия (шестое изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-397-51820-3 Полем
В 1964 году Dahlstrom и Fuxe (обсуждались в [2]), используя метод гистофлуоресценции Falck-Hillarp , наблюдали, что большинство серотонинергических сомов обнаруживаются в группах клеточных тела, которые ранее были обозначены как ядра Raphe.
- ^ Биндер М.Д., Хирокава Н. (2009). Энциклопедия нейробиологии . Берлин: Спрингер. п. 705. ISBN 978-3-540-23735-8 .
- ^ Группа ядерных ядер нейронов расположена вдоль ствола головного мозга от ярлыков « среднего мозга » до « oblongata », сосредоточенных на понах . ( См. Соответствующее изображение .)
- ^ Müller CP, Jacobs BL, Eds. (2009). Справочник по поведенческой нейробиологии серотонина (1 -е изд.). Лондон: академический. С. 51–59. ISBN 978-0-12-374634-4 .
- ^ AD, Herculano AM, Oliveira KR, Lima SM, Rocha FA, Freitas HR, et al. (Octber 2019). «Регуляция серотононской системы каинатом в птичьей сетчатке». Клеточная и молекулярная нейробиология . 39 (7): 1039–1049. Doi : 10.1007/s10571-019-00701-8 . PMID 31197744 . S2CID 189763144 .
- ^ Соммер С. (2009). «Серотонин в боли и боли в контроле». В Müller CP, Jacobs BL (Eds.). Справочник по поведенческой нейробиологии серотонина (1 -е изд.). Лондон: академический. С. 457–460. ISBN 978-0-12-374634-4 .
- ^ Хенслер Дж.Г. (2009). «Серотонин в режиме и эмоциях». В Müller CP, Jacobs BL (Eds.). Справочник по поведенческой нейробиологии серотонина (1 -е изд.). Лондон: академический. С. 367–399. ISBN 978-0-12-374634-4 .
- ^ Эндрюс П.В., Бхарвани А., Ли К.Р., Фокс М., Томсон Дж.А. (апрель 2015). «Является ли серотонин верхним или вниз? Эволюция серотонинергической системы и его роль в депрессии и антидепрессантов». Нейробиологии и биобиоэвиральные обзоры . 51 : 164–188. doi : 10.1016/j.neubiorev.2015.01.018 . PMID 25625874 . S2CID 23980182 .
- ^ Stahl SM, Mignon L, Meyer JM (март 2009 г.). «Что на первом месте: атипичное антипсихотическое лечение или кардиометаболический риск?» Полем Acta Psychiatrica Scandinavica . 119 (3): 171–179. doi : 10.1111/j.1600-0447.2008.01334.x . PMID 19178394 . S2CID 24035040 .
- ^ Buckland PR, Hoogendoorn B, Guy CA, Smith SK, Coleman SL, O'Donovan MC (март 2005 г.). «Низкая экспрессия генов, обеспечиваемой ассоциацией аллеля гена рецептора 5-HT2C с усилением веса, вызванным антипсихотическим средством». Американский журнал психиатрии . 162 (3): 613–615. doi : 10.1176/appi.ajp.162.3.613 . PMID 15741483 .
- ^ Холмс М.К., Французский К.Л., Секл младший (июнь 1997 г.). «Дисгуляция суточных ритмов серотонина 5-HT2C и экспрессии гена кортикостероидного рецептора в гиппокампе с ограничением пищи и глюкокортикоидами» . Журнал нейробиологии . 17 (11): 4056–4065. doi : 10.1523/jneurosci.17-11-04056.1997 . PMC 6573558 . PMID 9151722 .
- ^ Лейбовиц С.Ф. (1990). «Роль серотонина в расстройствах пищевого поведения». Наркотики . 39 (Suppl 3): 33–48. doi : 10.2165/00003495-199000393-00005 . PMID 2197074 . S2CID 8612545 .
- ^ McGuire, Michael (2013) «Вера, нейробиология фантазий, страхов и признаков» (Prometius Books)
- ^ Caspi N, Modai I, Barak P, Waisbourd A, Zbarsky H, Hirschmann S, et al. (Март 2001 г.). «Увеличение пиндолола у агрессивных пациентов с шизофренией: двойное слепое рандомизированное исследование кроссовера». Международная клиническая психофармакология . 16 (2): 111–115. doi : 10.1097/00004850-200103000-00006 . PMID 11236069 . S2CID 24822810 .
- ^ Это Z, Aizawa I, Takeuchi M, Tabe M, окрашен T (декабрь 1975 г.). Экскональная сила. Nihon Heikatsin Gakkaai Zasshi 11 (4): 244–246. PMID 1232434 .
- ^ Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, et al. (Ноябрь 1996). «Ассоциация связанных с тревогой признаками с полиморфизмом в регуляторной области транспортера серотонина». Наука . 274 (5292): 1527–1531. Bibcode : 1996sci ... 274.1527L . doi : 10.1126/science.274.5292.1527 . PMID 8929413 . S2CID 35503987 .
- ^ Битти Д.Т., Смит Дж.А. (май 2008 г.). «Фармакология серотонина в желудочно -кишечном тракте: обзор». Архив фармакологии Наунин-Шмидеберга . 377 (3): 181–203. doi : 10.1007/s00210-008-0276-9 . PMID 18398601 . S2CID 32820765 .
- ^ Rang HP (2003). Фармакология . Эдинбург: Черчилль Ливингстон. п. 187. ISBN 978-0-443-07145-4 .
- ^ De Wit R, Aapro M, Blower PR (сентябрь 2005 г.). «Существует ли фармакологическая основа различий в эффективности антагониста 5-HT3-рецептора у рефрактерных пациентов?». Химиотерапия рака и фармакология . 56 (3): 231–238. doi : 10.1007/s00280-005-1033-0 . PMID 15838653 . S2CID 27576150 .
- ^ Jump up to: а беременный Lauweryns JM, Cokelaere J, TheUnynck P (апрель 1973 г.). «Серотонин, продуцирующие нейроэпителиальные тела в кроличьей дыхательной слизистой оболочке». Наука . 180 (4084): 410–413. Bibcode : 1973sci ... 180..410L . doi : 10.1126/science.180.4084.410 . PMID 4121716 . S2CID 2809307 .
- ^ Пастор Л.М., Балеста Дж., Перес-Томас Р., Марин Дж.А., Эрнандес Ф., Мадрид Дж.Ф. (июнь 1987 г.). «Иммуноцитохимическая локализация серотонина в легком рептилий». Клеточные и тканевые исследования . 248 (3). Springer Science and Business Media LLC: 713–715. doi : 10.1007/bf00216504 . PMID 3301000 . S2CID 9871728 .
- ^ Jump up to: а беременный Sonstegard KS, Mailman RB, Cheek JM, Tomlin TE, Diagustine RP (ноябрь 1982). «Морфологическая и цитохимическая характеристика нейроэпителиальных тел в легких плода кролика. I. Исследования изолированных нейроэпителиальных тел». Экспериментальные исследования легких . 3 (3–4): 349–377. doi : 10.3109/01902148209069663 . PMID 6132813 .
- ^ Чанг В., Канда Х., Икеда Р., Лин Дж., Деберри Дж.Дж., Гу Дж. Г. (сентябрь 2016 г.). «Merkel Disc является серотонинергическим синапсом в эпидермисе для передачи тактильных сигналов у млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (37): E5491 - E5500. BIBCODE : 2016PNAS..113E5491C . doi : 10.1073/pnas.1610176113 . PMC 5027443 . PMID 27573850 .
- ^ Frost M, Andersen TE, Yadav V, Brixen K, Karsenty G, Kassem M (март 2010 г.). «Пациенты с фенотипом с высокой костью из-за мутации LRP5-T253I имеют низкие уровни серотонина в плазме» . Журнал исследований костей и минералов . 25 (3): 673–675. doi : 10.1002/jbmr.44 . PMID 20200960 . S2CID 24280062 .
- ^ Розен CJ (февраль 2009 г.). «Внесение в косную биологию: секреты серотонина». Природная медицина . 15 (2): 145–146. doi : 10.1038/nm0209-145 . PMID 19197289 . S2CID 5489589 .
- ^ Mödder UI, Achenbach SJ, Amin S, Riggs BL, Melton LJ, Khosla S (февраль 2010 г.). «Соотношение уровней серотонина в сыворотке к плотности кости и структурным параметрам у женщин» . Журнал исследований костей и минералов . 25 (2): 415–422. doi : 10.1359/jbmr.090721 . PMC 3153390 . PMID 19594297 .
- ^ Frost M, Andersen T, Gossiel F, Hansen S, Bollerslev J, Van Hul W, et al. (Август 2011 г.). «Уровни серотонина, склеростина, маркеров оборота костей, а также плотность костей и микроархитектуры у пациентов с фенотипом с высокой костью из-за мутации в LRP5» . Журнал исследований костей и минералов . 26 (8): 1721–1728. doi : 10.1002/jbmr.376 . PMID 21351148 .
- ^ Kode A, Mosialou I, Silva BC, Rached MT, Zhou B, Wang J, et al. (Октябрь 2012 г.). «Foxo1 организует функцию подавления костей серотонина, полученного из кишечника» . Журнал клинических исследований . 122 (10): 3490–3503. doi : 10.1172/jci64906 . PMC 3461930 . PMID 22945629 .
- ^ Ядав В.К., Баладжи С., Суреш П.С., Лю Хс, Лу Х, Ли З. и др. (Март 2010 г.). «Фармакологическое ингибирование синтеза серотонина, полученного из кишечника, является потенциальным анаболическим лечением кости при остеопорозе» . Природная медицина . 16 (3): 308–312. doi : 10.1038/nm.2098 . PMC 2836724 . PMID 20139991 .
- ^ Sugisaw E, Takayama Y, Kemmura N, Condo T, Hangyama S, Kugai Y, et al. (Август 2020). «Ощущение РНК с помощью кишечного пьезо1 необходимо для системного синтеза серотонина » Ячейка 182 (3): 609–624.e2 Doi : 10.1016/ j.cell.2020.06.0 PMID 32640190
- ^ Озанн С.Е., Хейлз С.Н. (январь 2004 г.). «Срок службы: догоняющий рост и ожирение у мышей-мужчин». Природа . 427 (6973): 411–412. Bibcode : 2004natur.427..411o . doi : 10.1038/427411b . PMID 14749819 . S2CID 40256021 .
- ^ CA, McGill CA, McGll HC, CD, Masuro 1986). "И не . Журнал клинических исследований 78 (4): 899–905. doi : 10.1172/ jci1 PMC 423712 . PMID 3760191 .
- ^ Хан П (июль 1984 г.). «Влияние размера мусора на холестерин в плазме и инсулин, а также некоторые ферменты печени и жировой ткани у взрослых грызунов». Журнал питания . 114 (7): 1231–1234. doi : 10.1093/JN/114.7.1231 . PMID 6376732 .
- ^ Попа Д., Лена С., Александр С., Адриен Дж (апрель 2008 г.). «Прочный синдром депрессии, вызванный снижением поглощения серотонина во время постнатального развития: данные из -за сна, стресса и поведения» . Журнал нейробиологии . 28 (14): 3546–3554. doi : 10.1523/jneurosci.4006-07.2008 . PMC 6671102 . PMID 18385313 .
- ^ Maciag D, Simpson KL, Coppinger D, Lu Y, Wang Y, Lin RC, et al. (Январь 2006 г.). «Воздействие неонатальных антидепрессантов оказывает длительное влияние на поведение и серотониновую схему» . Нейропсихофармакология . 31 (1): 47–57. doi : 10.1038/sj.npp.1300823 . PMC 3118509 . PMID 16012532 .
- ^ Maciag D, Williams L, Coppinger D, Paul IA (февраль 2006 г.). «Воздействие неонатального циталопрама вызывает длительные изменения в поведении, которые обращаются вспять при лечении взрослым имипрамином» . Европейский журнал фармакологии . 532 (3): 265–269. doi : 10.1016/j.ejphar.2005.12.081 . PMC 2921633 . PMID 16483567 .
- ^ Холден С (октябрь 2004 г.). «Нейробиология. Прозак лечение новорожденных мышей повышает беспокойство» . Наука . 306 (5697): 792. doi : 10.1126/science.306.5697.792 . PMID 15514122 .
- ^ Ansorge MS, Zhou M, Lira A, Hen R, Gingrich JA (октябрь 2004 г.). «Ранняя блокада 5-HT Transporter изменяет эмоциональное поведение у взрослых мышей» . Наука . 306 (5697): 879–881. Bibcode : 2004sci ... 306..879a . doi : 10.1126/science.1101678 . PMID 15514160 .
- ^ Lesurtel M, Graf R, Aleil B, Walther DJ, Tian Y, Jochum W, et al. (Апрель 2006 г.). «Сторонный серотонин опосредует регенерацию печени». Наука . 312 (5770): 104–107. Bibcode : 2006sci ... 312..104L . doi : 10.1126/science.1123844 . PMID 16601191 . S2CID 43189753 .
- ^ Matondo RB, Punt C, Homberg J, Toussaint MJ, Kisjes R, Korporaal SJ, et al. (Апрель 2009 г.). «Удаление транспортера серотонина у крыс нарушает гомеостаз серотонина без нарушения регенерации печени» . Американский журнал физиологии. Желудочно -кишечная физиология и печени . 296 (4): G963 - G968. doi : 10.1152/ajpgi.90709.2008 . PMID 19246633 . Архивировано из оригинала 28 декабря 2019 года . Получено 5 декабря 2019 года .
- ^ Collet C, Schiltz C, Geoffroy V, Maroteaux L, Launay JM, De Vernejoul MC (февраль 2008 г.). «Рецептор серотонина 5-HT2B контролирует костную массу посредством рекрутирования и пролиферации остеобластов» . FASEB Journal . 22 (2): 418–427. doi : 10.1096/fj.07-9209com . PMC 5409955 . PMID 17846081 .
- ^ Ядав В.К., Рю Дж.Х., Суда Н., Танака К.Ф., Гингрич Дж.А., Шюц Г. и др. (Ноябрь 2008 г.). «LRP5 контролирует образование кости, ингибируя синтез серотонина в двенадцатиперстной кишке» . Клетка . 135 (5): 825–837. doi : 10.1016/j.cell.2008.09.059 . PMC 2614332 . PMID 19041748 .
- «Ученые открывают для строительства кости, чтобы построить кости» . ScienceDaily (пресс -релиз). 1 декабря 2008 года.
- ^ McDuffie JE, Motley ED, Limbird LE, Maleque MA (март 2000 г.). «5-гидрокситриптамина стимулирует фосфорилирование митогеновой активации митогенов P44/P42 в культурах эндотелиальных клеток аорты бычьей аорты» . Журнал сердечно -сосудистой фармакологии . 35 (3): 398–402. doi : 10.1097/00005344-200003000-00008 . PMID 10710124 .
- ^ Noguchi M, Furukawa KT, Morimoto M (декабрь 2020 г.). «Легочные нейроэндокринные клетки: физиология, гомеостаз ткани и болезнь» . Модели заболевания и механизмы . 13 (12): DMM046920. doi : 10.1242/dmm.046920 . PMC 7774893 . PMID 33355253 .
- ^ Fuller RW (1980). «Фармакология центральных серотониновых нейронов». Ежегодный обзор фармакологии и токсикологии . 20 : 111–127. doi : 10.1146/annurev.pa.20.040180.000551 . PMID 6992697 .
- ^ Titeler M, Lyon RA, Glennon RA (1988). «Данные, связывающие радиолиганд, означают рецептор 5-HT2 мозга как место действия действий для LSD и фенилизопропиламина галлюциногенов». Психофармакология . 94 (2): 213–216. doi : 10.1007/bf00176847 . PMID 3127847 . S2CID 24179554 .
- ^ Николс де (2000). «Роль серотонинергических нейронов и 5-HT-рецепторов в действии галлюциногенов». В Baumgarten Hg, Gothert M (Eds.). Серотонинергические нейроны и 5-HT-рецепторы в ЦНС . Санта-Клара, Калифорния: Springer-Verlag Telos. ISBN 978-3-540-66715-5 .
- ^ Капур С., Симан П. (2002). «Антагонисты NMDA-рецептора кетамин и PCP оказывают прямое влияние на рецепторов дофамин D (2) и серотонина 5-HT (2)-рецепторов для моделей шизофрении» . Молекулярная психиатрия . 7 (8): 837–844. doi : 10.1038/sj.mp.4001093 . PMID 12232776 .
- ^ Депутат Джонсона, Хоффман А.Дж., Николс де (декабрь 1986 г.). «Влияние энантиомеров MDA, MDMA и связанных аналогов на [3H] серотонина и [3H] высвобождение дофамина из суперфузированных срезов мозга крысы». Европейский журнал фармакологии . 132 (2–3): 269–276. doi : 10.1016/0014-2999 (86) 90615-1 . PMID 2880735 .
- ^ Гудман Л.С., Брунтон Л.Л., Чабнер Б., Ноллманн Б.К. (2001). Фармакологическая основа терапии Гудмана и Гилмана . Нью-Йорк: МакГроу-Хилл. С. 459–461. ISBN 978-0-07-162442-8 .
- ^ Benmansour S, Cecchi M, Morilak DA, Gerhardt GA, Javors MA, Gould GG, et al. (Декабрь 1999). «Влияние хронических антидепрессантов на функцию транспортера серотонина, плотность и уровень мРНК» . Журнал нейробиологии . 19 (23): 10494–10501. doi : 10.1523/jneurosci.19-23-10494.1999 . PMC 6782424 . PMID 10575045 .
- ^ Бейчман Дж. Х., Балдассарра Л., Мик Х., Де Лука В., Кинг Н., Бендер Д. и др. (Июнь 2006 г.). «Полиморфизмы транспортера серотонина и постоянная, распространенная детская агрессия». Американский журнал психиатрии . 163 (6): 1103–1105. doi : 10.1176/appi.ajp.163.6.1103 . PMID 16741214 .
- ^ Pezawas L, Meyer-Lindenberg A, Drabant EM, Verchinski BA, Munoz KE, Kolachana BS, et al. (Июнь 2005 г.). «Полиморфизм 5-HTTLPR влияет на взаимодействие по поясной рамке человека-амидала: генетический механизм восприимчивости к депрессии». Nature Neuroscience . 8 (6): 828–834. doi : 10.1038/nn1463 . PMID 15880108 . S2CID 1864631 .
- ^ Schinka JA, Busch RM, Robichaux-Keene N (февраль 2004 г.). «Мета-анализ связи между полиморфизмом гена серотонина (5-HTTLPR) и тревожностью признаков» . Молекулярная психиатрия . 9 (2): 197–202. doi : 10.1038/sj.mp.4001405 . PMID 14966478 .
- ^ Jump up to: а беременный New Am, Nelson S, Leung JG (1 октября 2015 г.). Александр E, Susla Gm (ред.). «Психиатрические чрезвычайные ситуации в отделении интенсивной терапии». AACN Advanced Critical Care . 26 (4): 285–293, тест 294–295. doi : 10.4037/nci.0000000000000104 . PMID 26484986 .
- ^ Isbister GK, Bowe SJ, Dawson A, Whyte IM (2004). «Относительная токсичность селективных ингибиторов обратного захвата серотонина (СИОЗС) в передозировке». Журнал токсикологии. Клиническая токсикология . 42 (3): 277–285. doi : 10.1081/clt-120037428 . PMID 15362595 . S2CID 43121327 .
- ^ Данкли Э.Дж., Исбистер Г.К., Сиббритт Д., Доусон А.Х., Уайт Им (сентябрь 2003 г.). «Критерии токсичности серотонина охотника: простые и точные правила диагностического решения для токсичности серотонина» . QJM . 96 (9): 635–642. doi : 10.1093/qjmed/hcg109 . PMID 12925718 .
- ^ Фрэнк С (июль 2008 г.). «Распознавание и лечение синдрома серотонина» . Канадский семейный врач . 54 (7): 988–992. PMC 2464814 . PMID 186258822 .
- ^ Бойер Э.В., Шеннон М (март 2005 г.). «Синдром серотонина». Новая Англия Журнал медицины . 352 (11): 1112–1120. doi : 10.1056/nejmra041867 . PMID 15784664 .
- ^ Jump up to: а беременный Баскин С.И. (1991). Принципы сердечной токсикологии . Boca Raton: CRC Press. ISBN 978-0-8493-8809-5 Полем Получено 3 февраля 2010 года .
- ^ Jähnichen S, Horowski R, Pertz H. «Перголид и каберголин, но не лисурид демонстрируют агонистскую эффективность в серотонина 5-HT 2B рецепторах » (PDF) . Получено 3 февраля 2010 года .
- ^ Консультативный комитет по реакции на реакцию на наркотики, Австралия (2004). «Сердечная вальвулопатия с перголидом» . AUST AD ADR NURGACT React Bull . 23 (4). Архивировано из оригинала 27 июня 2012 года.
- ^ Schade R, Andersohn F, Suissa S, Haverkamp W, Garbe E (январь 2007 г.). «Агонисты дофамина и риск регургитации сердца» . Новая Англия Журнал медицины . 356 (1): 29–38. doi : 10.1056/nejmoa062222 . PMID 17202453 .
- ^ Zanettini R, Antonini A, Gatto G, Gentile R, Tesei S, Pezzoli G (январь 2007 г.). «Кластная болезнь сердца и использование дофаминовых агонистов для болезни Паркинсона» . Новая Англия Журнал медицины . 356 (1): 39–46. doi : 10.1056/nejmoa054830 . PMID 17202454 .
- ^ «Консультации по общественному здравоохранению по контролю за продуктами и лекарствами» . Управление по контролю за продуктами и лекарствами . 29 марта 2007 г. Получено 7 февраля 2010 года .
- ^ «MedWatch - 2007 г. Предостатки информации о безопасности. Permax (Pergolide) и общие эквиваленты» . США Управление по контролю за продуктами и лекарствами . 29 марта 2007 г. Получено 30 марта 2007 года .
- ^ Тайлер В.Е. (сентябрь 1958 г.). «Появление серотонина в галлюциногенном грибах». Наука . 128 (3326): 718. Bibcode : 1958sci ... 128..718t . doi : 10.1126/science.128.3326.718 . PMID 13580242 .
- ^ Джонсон DJ, Сандерсон H, Brain RA, Wilson CJ, Solomon KR (май 2007). «Токсичность и опасность селективного серотонина обратного ингибитора антидепрессантов флуоксетина, флувоксамина и сертралина к водорослям». Экотоксикология и безопасность окружающей среды . 67 (1): 128–139. Bibcode : 2007ecoes..67..128j . doi : 10.1016/j.ecoenv.2006.03.016 . PMID 16753215 .
- ^ McGowan K, Guerina V, Wicks J, Donowitz M (1985). «Секреторные гормоны Энтомебы Histolytica ». Симпозиум Фонда CIBA . Симпозии Фонда Новартиса. Тол. 112. С. 139–154. doi : 10.1002/9780470720936.CH8 . ISBN 978-0-470-72093-6 Полем PMID 2861068 .
- ^ Бану Н., Заиди К.Р., Мехди Г., Мансур Т (июль 2005 г.). «Нейрогуморальные изменения и их роль в амебиазе» . Индийский журнал клинической биохимии . 20 (2): 142–145. doi : 10.1007/bf02867414 . PMC 3453840 . PMID 23105547 .
- ^ Acharya DP, Sen Mr, Sen PC (август 1989 г.). «Влияние экзогенного 5-гидрокситриптамина на патогенность entamoeba histolytica у экспериментальных животных». Индийский журнал экспериментальной биологии . 27 (8): 718–720. PMID 2561282 .
- ^ Schröder P, Abele C, Gohr P, Stuhlfauth-Roisch U, Grosse W (1999). «Последние об фермере серотонина биосинтеза в семенах грецкого ореха». Триптофан, серотонин и мелатонин . Достижения в области экспериментальной медицины и биологии. Тол. 467. С. 637–644. doi : 10.1007/978-1-4615-4709-9_81 . ISBN 978-0-306-46204-7 Полем PMID 10721112 .
- ^ Pénez N, Culioli G, Pérez T, Briand JF, Thomas OP, Blache Y (октябрь 2011 г.). «Противоречивые свойства простых индольных и пуриновых алкалоидов из средиземноморской горгонианской парамаричея clavata». Журнал натуральных продуктов . 74 (10): 2304–2308. doi : 10.1021/np200537v . PMID 21939218 .
- ^ Guillén-Casla V, Rosales-Conrado N, León-González ME, Pérez-Arribas LV, Polo-Díez LM (апрель 2012 г.). «Определение серотонина и его предшественников в образцах шоколада с помощью капиллярной жидкой хроматографии с обнаружением масс -спектрометрии». Журнал хроматографии а . 1232 : 158–165. doi : 10.1016/j.chroma.2011.11.037 . PMID 22186492 .
- ^ Pelagio-Flores R, Ortíz-Castro R, Méndez-Bravo A, Macías-Rodríguez L, López-Bucio J (март 2011 г.). «Серотонин, полученный из триптофана сигнал, консервативный у растений и животных, регулирует архитектуру корневой системы, вероятно, действует как естественный ингибитор ауксина в арабидопсисе Thaliana» . Физиология растений и клеток . 52 (3). Издательство Оксфордского университета (OUP): 490–508. doi : 10.1093/pcp/pcr006 . PMID 21252298 .
- ^ Jump up to: а беременный Powell JJ, Carere J, Fitzgerald TL, Stiller J, Covarelli L, Xu Q, et al. (Март 2017). «Фузариум -гнильный патоген фузариум псевдограммия запускает набор транскрипционных и метаболических изменений в хлебной пшенице (Triticum aestivum L.)» . Анналы ботаники . 119 (5). Издательство Оксфордского университета (OUP): 853–867. doi : 10.1093/aob/mcw207 . PMC 5604588 . PMID 27941094 . S2CID 3823345 .
- ^ Du Fall LA, Solomon PS (октябрь 2013 г.). «Некротрофный эффектор SNTOXA индуцирует синтез нового фитоалексина в пшенице» . Новый фитолог . 200 (1). Уайли : 185–200. doi : 10.1111/nph.12356 . PMID 23782173 .
- ^ Pasquet JC, Chaouch S, Macadré C, Balzergue S, Huguet S, Martin-Magniette ML, et al. (Июль 2014). «Дифференциальная экспрессия генов и метаболомический анализ брахиподий-дистахиона, инфицированных дезоксиниваленолом, продуцирующими и не продуцирующими штаммами Fusarium graminearum» . BMC Genomics . 15 (1). Biomed Central : 629. DOI : 10.1186/1471-2164-15-629 . PMC 4124148 . PMID 25063396 .
- ^ Zeiss DR, Piater LA, Dubery IA (февраль 2021 г.). «Амиды гидроксициннамата: интригующие конъюгаты защитных метаболитов растений». Тенденции в науке о растениях . 26 (2). Клетовая пресса : 184–195. Бибкод : 2021tps .... 26..184Z . doi : 10.1016/j.tlants.2020.09.011 . PMID 33036915 . S2CID 222256660 .
- ^ Jonz MG, Ekaterinimercier A, Joffrepotter JW (2001). «Влияние 5-HT (серотонина) на репродуктивное поведение у Heterodera Schachtii (Nematoda)». Канадский журнал зоологии . 79 (9): 1727. DOI : 10.1139/Z01-135 .
- ^ Sawin er, Ranganathan R, Horvitz HR (июнь 2000 г.). «C. Elegans Locomotory Sceent модулируется окружающей средой посредством дофаминергического пути и опытом через серотонинергический путь» . Нейрон . 26 (3): 619–631. doi : 10.1016/s0896-6273 (00) 81199-x . PMID 10896158 . S2CID 9247380 .
- ^ Niacaris T, Avery L (январь 2003 г.). «Серотонин регулирует реполяризацию мышцы глотки C. elegans» . Журнал экспериментальной биологии . 206 (Pt 2): 223–231. doi : 10.1242/jeb.00101 . PMC 4441752 . PMID 12477893 .
- ^ Россо М.Н., Джонс Дж. Т., Абад П. (2009). «РНКи и функциональная геномика у паразитических нематод растений». Ежегодный обзор фитопатологии . 47 (1). Ежегодные обзоры : 207–232. doi : 10.1146/annurev.phyto.112408.132605 . PMID 19400649 . п. 218:
и серотонин регулируют активность нейронов М3, которые прямое сокращение глотки во время C. elegans питающих осьупамин , Экспрессируется в кишечнике и пищеводах.
- ^ Yeh Sr, Fricke RA, Edwards DH (январь 1996 г.). «Влияние социального опыта на серотонинергическую модуляцию схемы побега раков». Наука . 271 (5247): 366–369. Bibcode : 1996sci ... 271..366y . Citeseerx 10.1.1.470.6528 . doi : 10.1126/science.271.5247.366 . PMID 8553075 . S2CID 1575533 .
- ^ Fry Bg, Roelants K, Champagne DE, Scheib H, Tyndall JD, King GF, et al. (2009). «Токсикогеномная мультивселенная: сходящаяся рекрутирование белков в яды животных» . Ежегодный обзор геномики и генетики человека . 10 (1). Ежегодные обзоры : 483–511. doi : 10.1146/annurev.genom.9.081307.164356 . PMID 19640225 .
- ^ «Серотонин, рецепторы серотонина и их действия у насекомых» . Нейротрансмиттер . 2 : 1–14. 2015. DOI : 10.14800/nt.314 .
- ^ Jump up to: а беременный в Schoofs L, De Loof A, Van Hiel MB (январь 2017 г.). «Нейропептиды как регуляторы поведения у насекомых» . Ежегодный обзор энтомологии . 62 (1). Ежегодные обзоры : 35–52. doi : 10.1146/annurev-ento-031616-035500 . PMID 27813667 .
- ^ Jump up to: а беременный Ван Х, Кан Л (7 января 2014 г.). «Молекулярные механизмы изменения фазы в саранчи» . Ежегодный обзор энтомологии . 59 (1). Ежегодные обзоры : 225–244. doi : 10.1146/annurev-ento-011613-162019 . PMID 24160426 .
п. 231,
Изменение количества нескольких потенциальных нейротрансмиттеров ... таких как серотонин ... может играть важную роль в ремоделировании ЦНС во время фазового изменения (26, 56, 80).
п. 233,
В саранчке S. gregaria количество серотонина в грудных ганглиях было положительно коррелировано с степени общительного поведения, вызванного различными периодами толпы. Ряд фармакологических и поведенческих экспериментов показал, что серотонин играет ключевую роль в индукции начальной поведенческой грегаризации (2, 80). Тем не менее, серотонин не несет ответственности за поддержание общительного поведения, потому что его количество в долгосрочной общительной саранчве меньше, чем вдвое, в долгосрочной солидарной саранчке (80). В L. Migratoria инъекция серотонина также может немного инициировать общительное поведение, но серотонин, когда сопровождающий толпу, вызвала более солидарное поведение, чем инъекция серотонина в одиночку (48). Значительные различия в уровнях серотонина не были обнаружены в тканях головного мозга между двумя фазами L. Migratoria . Недавний отчет Tanaka & Nishide (97) измерял поведение притяжения/избегания у S. gregaria после одиночных и множественных инъекций серотонина в разных концентрациях. Серотонин оказал лишь кратковременное влияние на уровень некоторых локомоторных действий и не участвовал в контроле общительного поведения (97). Кроме того, неясно, как нейротрансмиттер влияет на это уникальное поведение, потому что модель бинарной логистической регрессии, используемая в этих исследованиях для поведенческого анализа, сфокусированной в основном только на одном поведенческом параметре, представляющем общее состояние фазы. Очевидно, что изменение поведенческой фазы может включать альтернативные регуляторные механизмы у разных видов саранчи. Следовательно, эти исследования показывают, что регуляторные механизмы ЦНС, регулирующие инициацию и поддержание фазового изменения, являются специфичными для видов и включают взаимодействие между этими нейротрансмиттерами.
Учитывая ключевые роли аминергической передачи сигналов, каковы пути ниже по течению, связанные с установлением долговременной памяти? Ott et al. (63) исследовали роль [] протеинкиназы [] в фазовом изменении в S. gregaria : ... CAMP-зависимая протеинкиназа A (PKA). Благодаря использованию фармакологического и RNAI вмешательства эти авторы продемонстрировали, что PKA ... играет важную роль в модулировании склонности саранчи к приобретению и выражению общительного поведения. ... К сожалению, хотя корреляция между серотонином и PKA была выдвинута гипотетической, прямые доказательства не были предоставлены. - ^ Jump up to: а беременный Zhang L, Lecoq M, Latchininsky A, Hunter D (январь 2019). «Саранча и управление кузнечиком» . Ежегодный обзор энтомологии . 64 (1). Ежегодные обзоры : 15–34. doi : 10.1146/annurev-ento-011118-112500 . PMID 30256665 . S2CID 52843907 . п. 20:
... Грегаризация вызывает ... тактильную стимуляцию ... тактильные стимулы запускают увеличение биогенных аминов, особенно серотонина, в нервной системе саранчи (1, 116); Эти амины играют критическую роль в нейрофизиологии изменения поведенческой фазы саранчи.
- ^ Jump up to: а беременный Anstey ML, Rogers SM, Ott Sr, Burrows M, Simpson SJ (январь 2009 г.). «Серотонин опосредует поведенческую грамеревацию, лежащую в основе образования роя в пустынной саранчке». Наука . 323 (5914): 627–630. Bibcode : 2009Sci ... 323..627a . doi : 10.1126/science.1165939 . PMID 19179529 . S2CID 5448884 .
- Морган Дж (29 января 2009 г.). «Саранча рой» «Высоте» на серотонине » . BBC News .
- ^ Sitaraman D, Laferriere H, Birman S, Zars T (июнь 2012 г.). «Серотонин имеет решающее значение для вознагражденной обонятельной кратковременной памяти у дрозофилы». Журнал нейрогенетики . 26 (2): 238–244. doi : 10.3109/01677063.2012.666298 . PMID 22436011 . S2CID 23639918 .
- ^ Bicker G, Menzel R (январь 1989 г.). «Химические коды для контроля поведения в членистоногих». Природа . 337 (6202): 33–39. Bibcode : 1989natur.337 ... 33b . doi : 10.1038/337033a0 . PMID 2562906 . S2CID 223750 .
- ^ Cai M, Li Z, Fan F, Huang Q, Shao X, Song G (март 2010 г.). «Дизайн и синтез новых инсектицидов, основанных на серотонинергическом лиганде 1-[(4-аминофенил) этил] -4- [3- (трифторметил) фенил] пиперазин (PAPP)». Журнал сельскохозяйственной и пищевой химии . 58 (5): 2624–2629. doi : 10.1021/jf902640u . PMID 20000410 .
- ^ Манахан С.Е. (2002). Токсикологическая химия и биохимия (3 -е изд.). CRC Press. п. 393. ISBN 978-1-4200-3212-3 .
- ^ Postma TL (2009). «Нейротоксические яды и яды» . В Dobbs MR (ed.). Клиническая нейротоксикология . WB Saunders. С. 463–489. doi : 10.1016/b978-032305260-3.50049-6 . ISBN 978-0-323-05260-3 .
- ^ Gadenne C, Barrozo RB, Anton S (11 марта 2016 г.). «Пластичность в одурчение насекомых: запах или не пахнуть?». Ежегодный обзор энтомологии . 61 (1). Ежегодные обзоры : 317–333. doi : 10.1146/annurev-ento-010715-023523 . HDL : 11336/19586 . PMID 26982441 . S2CID 207568844 .
- ^ Дирик Х.А., Гринспен Р.Дж. (май 2007 г.). «Серотонин и нейропептид F оказывают противоположное модуляторное воздействие на агрессию мухи». Природа генетика . 39 (5): 678–682. doi : 10.1038/ng2029 . PMID 17450142 . S2CID 33768246 .
- ^ Stoffolano JG , Haselton по адресу (7 января 2013 г.). «Взрослый урожай Dipteran: уникальный и пропущенный орган». Ежегодный обзор энтомологии . 58 (1). Ежегодные обзоры : 205–225. doi : 10.1146/annurev-ento-120811-153653 . PMID 23317042 .
- ^ Jump up to: а беременный в дюймовый и фон Bacqué-Cazenave J, Bharatiya R, Barrière G, Delbecque JP, Bouguiyoud N, Giovanni G, et al. (Февраля 2020). «Серотонин в познании и поведении животных» . Международный журнал молекулярных наук . 21 (5): 1649. doi : 10.3390/ijms21051649 . PMC 7084567 . PMID 32121267 .
- ^ Jump up to: а беременный в дюймовый и Lucki I (август 1998 г.). «Спектр поведения под влиянием серотонина» . Биологическая психиатрия . 44 (3): 151–162. doi : 10.1016/s0006-3223 (98) 00139-5 . PMID 9693387 . S2CID 3001666 .
- ^ Jump up to: а беременный в Backström T, Winberg S (25 октября 2017 г.). «Серотонин координирует ответы на социальное стресс, чему мы можем учиться у рыбы» . Границы в нейробиологии . 11 : 595. DOI : 10.3389/fnins.2017.00595 . PMC 5669303 . PMID 29163002 .
- ^ Бергер М., Грей Дж.А., Рот Б.Л. (1 февраля 2009 г.). «Расширенная биология серотонина» . Ежегодный обзор медицины . 60 (1): 355–366. doi : 10.1146/annurev.med.60.042307.110802 . PMC 5864293 . PMID 19630576 .
- ^ Alberghina D, Piccione G, Pumilia G, Gioè M, Rizzo M, Raffo P, et al. (Июль 2019). «Ежедневные колебания мочи серотонина и кортизола у здоровых укрытий собак и влияние внутривидового социального воздействия». Физиология и поведение . 206 : 1–6. doi : 10.1016/j.physbeh.2019.03.016 . PMID 30898540 . S2CID 81965422 .
- ^ Риггио Г., Марити С., Серги В., Диверио С., Газзано А (декабрь 2020 г.). «Концентрации серотонина и триптофана в сыворотке у приютов, демонстрирующих различные поведенческие реакции на потенциально стрессовую процедуру» . Ветеринарные науки . 8 (1): 1. DOI : 10.3390/vetsci8010001 . PMC 7824451 . PMID 33374183 .
- ^ Hydbring-Sandberg E, Larsson E, Modej A, Höglund OV (июль 2021 г.). «Краткосрочное влияние овариогистерэктомии на серотонин мочи, кортизол, тестостерон и прогестерон у сук» . BMC Research Notes . 14 (1): 265. doi : 10.1186/s13104-021-05680-y . PMC 8272283 . PMID 34246304 .
- ^ Jump up to: а беременный в Winberg S, Thörnqvist Po (июнь 2016 г.). «Роль серотонина мозга в модулировании поведения рыбы» . Текущая зоология . 62 (3): 317–323. doi : 10.1093/cz/zow037 . PMC 5804243 . PMID 29491919 .
- ^ Jump up to: а беременный в дюймовый Оливье Б., Мос Дж., Ван дер Хейден Дж., Хартог Дж. (1 февраля 1989 г.). «Серотонинергическая модуляция социальных взаимодействий у изолированных самцов мышей». Психофармакология . 97 (2): 154–156. doi : 10.1007/bf00442239 . PMID 2498921 . S2CID 37170174 .
- ^ Хубер Р., Смит К., Делаго А., Исакссон К., Кравитц Е.А. (май 1997). «Серотонин и агрессивная мотивация у ракообразных: изменение решения отступить» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (11): 5939–5942. Bibcode : 1997pnas ... 94.5939H . doi : 10.1073/pnas.94.11.5939 . PMC 20885 . PMID 9159179 .
- ^ Санчес К.Л., Бискуп С.С., Герперц С., Габер Т.Дж., Кун С.М., Худ Ш.Х. и др. (Май 2015). «Роль серотонина (5-HT) в поведенческом контроле: результаты исследований животных и клинических последствий» . Международный журнал нейропсихофармакологии . 18 (10): PYV050. doi : 10.1093/ijnp/pyv050 . PMC 4648158 . PMID 25991656 .
- ^ Jump up to: а беременный в дюймовый Петерсен К.Л., Херли Л.М. (октябрь 2017 г.). «Поместить его в контекст: связывание слуховой обработки с цепями социального поведения в мозге позвоночных» . Интегративная и сравнительная биология . 57 (4): 865–877. doi : 10.1093/icb/icx055 . PMC 6251620 . PMID 28985384 .
- ^ Loer CM, Kenyon CJ (декабрь 1993 г.). «Мутанты с дефицитом серотонина и мужское спаривание в нематоде Caenorhabditis elegans» . Журнал нейробиологии . 13 (12): 5407–5417. doi : 10.1523/jneurosci.13-12-05407.1993 . PMC 6576401 . PMID 8254383 .
- ^ Lipton J, Kleemann G, Ghosh R, Lints R, Emmons SW (август 2004 г.). «Мат поиск в Caenorhabditis elegans: генетическая модель для сексуального влечения в простом беспозвоночном» . Журнал нейробиологии . 24 (34): 7427–7434. doi : 10.1523/jneurosci.1746-04.2004 . PMC 6729642 . PMID 15329389 .
- ^ Jump up to: а беременный Мураками Х, Мураками С (август 2007 г.). «Рецепторы серотонина антагонистически модулируют долговечность Caenorhabditis elegans» . Старение ячейки . 6 (4): 483–488. doi : 10.1111/j.1474-9726.2007.00303.x . PMID 17559503 . S2CID 8345654 .
- ^ Kaplan DD, Zimmermann G, Suyama K, Meyer T, Scott MP (июль 2008 г.). «GTPase, семейство нуклеостеминов, NS3, действует в серотонинергических нейронах для регулирования передачи сигналов инсулина и контрольного размера тела» . Гены и развитие . 22 (14): 1877–1893. doi : 10.1101/gad.1670508 . PMC 2492735 . PMID 18628395 .
- ^ Ruaud AF, Thummel CS (июль 2008 г.). «Сигнализация серотонина и инсулина поднимается для контроля роста в дрозофиле» . Гены и развитие . 22 (14): 1851–1855. doi : 10.1101/gad.1700708 . PMC 2735276 . PMID 18628391 .
- ^ Дэвидсон С., Проконов Д., Талер М., Мааян Р., Харелл Д., Гил-Ад И. и др. (Февраль 2009 г.). «Влияние воздействия селективных ингибиторов обратного захвата серотонина в утробежном порядке на рост плода: потенциальная роль для осями IGF-I и HPA» . Педиатрическое исследование . 65 (2): 236–241. doi : 10.1203/pdr.0b013e318193594a . PMID 19262294 .
- ^ Бен Арос Дж., Лаффонт С., Чатенай Д (октябрь 2009 г.). Brezina V (ред.). «Молекулярная и сенсорная база поведения с двумя государствами, связанным с пищевой пищей у C. elegans» . Plos один . 4 (10): E7584. Bibcode : 2009ploso ... 4.7584b . doi : 10.1371/journal.pone.0007584 . PMC 2762077 . PMID 19851507 .
- ^ Sze JY, Victor M, Loer C, Shi Y, Rivkun G (февраль 2000 г.). «Пищевые и метаболические сигнальные дефекты в каендере мутанта Seronin Seronin-синтез» Природа 403 (6769): 560–5 Bibcode : 2000natur.403..560s Doi : 10.1038/ 35000609 10676966PMID 4394553S2CID
- ^ Мураками Х., Бессингер К., Хеллманн Дж., Мураками С. (июль 2008 г.). «Манипулирование сигналом серотонина подавляет раннюю фазу поведенческого старения у Caenorhabditis elegans». Нейробиология старения . 29 (7): 1093–1100. doi : 10.1016/j.neurobiolaging.2007.01.013 . PMID 17336425 . S2CID 37671716 .
- ^ Côté F, Thevenot E, Fligny C, Fromes Y, Darmon M, Ripoche MA, et al. (Ноябрь 2003). «Нарушение гена нениронального TPH1 демонстрирует важность периферического серотонина в функции сердца» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (23): 13525–13530. BIBCODE : 2003PNAS..10013525C . doi : 10.1073/pnas.2233056100 . PMC 263847 . PMID 14597720 .
- ^ Alarcón «Биотрансформация производных миоцелийскими культурами » Naturforschung c 63 (1–2): 82–84. два 10.1515/znc-2008-1-215: 18386493PMID 29472174S2CID
- ^ Sismaet HJ, Goluch Ed (июнь 2018 г.). «Электрохимические зонды поведения микробного сообщества» . Ежегодный обзор аналитической химии . 11 (1). Ежегодные обзоры : 441–461. Bibcode : 2018arac ... 11..441s . doi : 10.1146/annurev-anchem-061417-125627 . PMID 29490192 . S2CID 3632265 . п. 449:
Таблица 1 Соответствующие потенциальные пики для различных электроактивных биомолекул, которые производятся или потребляются микробами, сообщенными в литературе а ... серотонин | Оксид индия олова | +0,67 | 66
- ^ Хенсон Дж. М., Батлер М.Дж., Дэй А.В. (1999). «Темная сторона мицелия: меланины фитопатогенных грибов». Ежегодный обзор фитопатологии . 37 (1). Ежегодные обзоры : 447–471. doi : 10.1146/annurev.phyto.37.1.447 . PMID 11701831 .
- ^ Энтони М (декабрь 1984 г.). «Антагонисты серотонина». Австралийский и новозеландский журнал медицины . 14 (6): 888–895. doi : 10.1111/j.1445-5994.1984.tb03802.x . PMID 6398056 . S2CID 28327178 .
- ^ Erspamer V (декабрь 1954 г.). «Фармакология индоле-алкиламинов». Фармакологические обзоры . 6 (4): 425–487. PMID 13236482 .
- ^ Негри Л (2006). «[Vittorio Erspamer (1909-1999)]» . Лекарство на протяжении веков . 18 (1): 97–113. PMID 17526278 .
- ^ Rapport MM, Green AA, Page IH (декабрь 1948 г.). «Сывороточный вазоконстриктор, серотонин; изоляция и характеристика» . Журнал биологической химии . 176 (3): 1243–1251. doi : 10.1016/s0021-9258 (18) 57137-4 . PMID 18100415 .
- ^ Feldberg W, Toh CC (февраль 1953 г.). «Распределение 5-гидрокситриптамина (серотонин, энтехман) в стене пищеварительного тракта» . Журнал физиологии . 119 (2–3): 352–362. doi : 10.1113/jphysiol.1953.sp004850 . PMC 1392800 . PMID 13035756 .
- ^ Scifinder - Деталь вещества серотонина. Доступ (4 ноября 2012 г.). [ Полная цитата необходима ]
- ^ Twarog BM, Page IH (октябрь 1953 г.). «Содержание серотонина в некоторых тканях и моче млекопитающих и метод для его определения» . Американский журнал физиологии . 175 (1): 157–161. doi : 10.1152/ajplegacy.1953.175.1.157 . PMID 13114371 .
- ^ Страница IH (июль 1954 г.). «Серотонин (5-гидрокситриптамина)». Физиологические обзоры . 34 (3). Американское физиологическое общество : 563–588. doi : 10.1152/physrev.1954.34.3.563 . PMID 13185755 .
Дальнейшее чтение
[ редактировать ]- Gutknecht L, Jacob C, Strobel A, Kriegebaum C, Müller J, Zeng Y, et al. (Июнь 2007 г.). «Вариация гена триптофана гидроксилазы-2 влияет на личностные черты и расстройства, связанные с эмоциональной дисрегуляцией» . Международный журнал нейропсихофармакологии . 10 (3): 309–320. doi : 10.1017/s1461145706007437 . PMID 17176492 .
Внешние ссылки
[ редактировать ]- 5-гидрокситриптамина MS Spectrum
- Серотонин связан с белками в PDB
- PsychotropicalResearch Обширные обзоры серотонинергических препаратов и синдрома серотонина.
- Молекула месяца: серотонин в Университете Бристоля
- 60-секундная псих: не справедливо! Мой уровень серотонина низкий , научный американец
- Тест серотонина на Clinlab Navigator .