стр.53
| ТП53 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TP53 , BCC7, LFS1, P53, TRP53, опухолевый белок p53, BMFS5, гены, p53 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 191170 ; МГИ : 98834 ; Гомологен : 460 ; Генные карты : TP53 ; ОМА : TP53 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||





p53 , также известный как опухолевый белок P53 , клеточный опухолевый антиген p53 ( название UniProt ) или белок 53, связанный с трансформацией (TRP53), представляет собой регуляторный белок, который часто мутирует при раке человека. Белки p53 (первоначально считались и часто назывались одним белком) имеют решающее значение для позвоночных , поскольку они предотвращают образование рака . [ 5 ] Таким образом, р53 был описан как «хранитель генома » из-за его роли в сохранении стабильности путем предотвращения мутаций генома. [ 6 ] Отсюда TP53 [ примечание 1 ] классифицируется как ген-супрессор опухоли . [ 7 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ]
Ген TP53 является наиболее часто мутирующим геном (>50%) при раке человека, что указывает на то, что ген TP53 играет решающую роль в предотвращении образования рака. [ 5 ] Ген TP53 кодирует белки, которые связываются с ДНК и регулируют экспрессию генов, предотвращая мутации генома. [ 12 ] Помимо полноразмерного белка, ген TP53 человека кодирует по меньшей мере 12 изоформ белка . [ 13 ]
Ген
[ редактировать ]У человека ген TP53 расположен на коротком плече 17-й хромосомы (17p13.1). [ 7 ] [ 8 ] [ 9 ] [ 10 ] Ген занимает длину 20 т.п.н. , имеет некодирующий экзон 1 и очень длинный первый интрон длиной 10 т.п.н., перекрывающий ген Hp53int1 . Кодирующая последовательность содержит пять областей, демонстрирующих высокую степень консервативности у позвоночных, преимущественно в экзонах 2, 5, 6, 7 и 8, но последовательности, обнаруженные у беспозвоночных, демонстрируют лишь отдаленное сходство с TP53 млекопитающих. [ 14 ] TP53 ортологи [ 15 ] были идентифицированы у большинства млекопитающих , для которых доступны полные данные о геноме.
человека TP53 Ген
[ редактировать ]У людей распространенный полиморфизм включает замену аргинина на пролин в положении кодона 72 экзона 4. Многие исследования изучали генетическую связь между этой вариацией и предрасположенностью к раку; однако результаты оказались противоречивыми. Например, метаанализ 2009 года не смог выявить связь с раком шейки матки. [ 16 ] Исследование 2011 года показало, что мутация пролина TP53 действительно оказывает глубокое влияние на риск рака поджелудочной железы у мужчин. [ 17 ] Исследование арабских женщин показало, что гомозиготность пролина по кодону 72 TP53 связана со снижением риска рака молочной железы. [ 18 ] Одно исследование показало, что TP53 полиморфизмы кодона 72 , MDM2 SNP309 и A2164G могут в совокупности быть связаны с предрасположенностью к раку, не связанному с ротоглоткой, и что SNP309 MDM2 в сочетании с кодоном 72 TP53 может ускорять развитие рака, не связанного с ротоглоткой, у женщин. [ 19 ] Исследование 2011 года показало, что полиморфизм кодона 72 TP53 связан с повышенным риском рака легких. [ 20 ]
Мета-анализ 2011 года не выявил значимой связи между полиморфизмом кодона 72 TP53 и риском колоректального рака. [ 21 ] и риск рака эндометрия. [ 22 ] Исследование, проведенное в 2011 году среди бразильской когорты новорожденных, выявило связь между немутантным аргинином TP53 и людьми, не имеющими семейного анамнеза рака. [ 23 ] Другое исследование 2011 года показало, что гомозиготный (Pro/Pro) генотип p53 связан со значительно повышенным риском развития почечно-клеточного рака. [ 24 ]
Функция
[ редактировать ]Повреждение и восстановление ДНК
[ редактировать ]p53 играет роль в регуляции или продвижении клеточного цикла, апоптозе и стабильности генома посредством нескольких механизмов:
- Он может активировать белки репарации ДНК , когда ДНК повреждена. Таким образом, это может быть важным фактором старения . [ 25 ]
- Он может остановить рост, удерживая клеточный цикл в точке регуляции G1/S при распознавании повреждений ДНК. Если он удерживает клетку здесь достаточно долго, белки репарации ДНК успеют исправить повреждение, и клетка сможет продолжить свое существование. клеточный цикл.
- Он может инициировать апоптоз (т.е. запрограммированную гибель клеток ), если повреждение ДНК окажется непоправимым.
- Это важно для реакции старения коротких теломер .
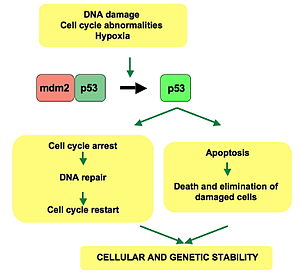
WAF1/CIP1 кодирует p21 и сотни других нижестоящих генов. p21 (WAF1) связывается с комплексами G1 - S / CDK ( CDK4 / CDK6 , CDK2 и CDK1 ) (молекулами, важными для перехода G1/S в клеточном цикле), ингибируя их активность.
Когда p21(WAF1) образует комплекс с CDK2, клетка не может перейти к следующему этапу клеточного деления. Мутантный р53 больше не будет эффективно связывать ДНК, и, как следствие, белок р21 не сможет выступать в качестве «стоп-сигнала» для деления клеток. [ 26 ] Исследования эмбриональных стволовых клеток человека (чЭСК) обычно описывают нефункциональную ось p53-p21 пути контрольной точки G1/S с последующим значением для регуляции клеточного цикла и реакции на повреждение ДНК (DDR). Важно отметить, что мРНК р21 явно присутствует и активируется после DDR в ЭСК, но белок р21 не обнаруживается. В этом типе клеток p53 активирует многочисленные микроРНК (например, miR-302a, miR-302b, miR-302c и miR-302d), которые непосредственно ингибируют экспрессию p21 в ЭСК.
Белок p21 напрямую связывается с комплексами циклин-CDK, которые ускоряют клеточный цикл и ингибируют их киназную активность, тем самым вызывая остановку клеточного цикла, позволяя осуществить восстановление. p21 может также опосредовать остановку роста, связанную с дифференцировкой, и более постоянную остановку роста, связанную с клеточным старением. Ген p21 содержит несколько элементов ответа p53, которые опосредуют прямое связывание белка p53, что приводит к активации транскрипции гена, кодирующего белок p21.
Пути p53 и RB1 связаны через p14ARF, что повышает вероятность того, что эти пути могут регулировать друг друга. [ 27 ]
Экспрессия p53 может стимулироваться ультрафиолетовым светом, который также вызывает повреждение ДНК. В этом случае р53 может инициировать события, ведущие к загару . [ 28 ] [ 29 ]
Стволовые клетки
[ редактировать ]Уровни р53 играют важную роль в поддержании стволовых клеток на протяжении всего развития и на протяжении всей жизни человека.
В эмбриональных стволовых клетках человека (чЭСК) р53 поддерживается на низких неактивных уровнях. [ 30 ] Это связано с тем, что активация р53 приводит к быстрой дифференцировке ЭСК. [ 31 ] Исследования показали, что нокаут р53 задерживает дифференцировку, а добавление р53 вызывает спонтанную дифференцировку, показывая, как р53 способствует дифференцировке ЭСК и играет ключевую роль в клеточном цикле в качестве регулятора дифференцировки. Когда р53 стабилизируется и активируется в ЭСК, он увеличивает р21, чтобы установить более длинный G1. Обычно это приводит к отмене входа в S-фазу, что останавливает клеточный цикл в G1, что приводит к дифференцировке. Однако работа с эмбриональными стволовыми клетками мышей недавно показала, что экспрессия P53 не обязательно приводит к дифференцировке. [ 32 ] p53 также активирует миР-34a и миР-145 , которые затем подавляют факторы плюрипотентности hESC, дополнительно стимулируя дифференцировку. [ 30 ]
Во взрослых стволовых клетках регуляция р53 важна для поддержания стволовости в нишах взрослых стволовых клеток . Механические сигналы, такие как гипоксия, влияют на уровни p53 в этих клетках ниши посредством факторов, индуцируемых гипоксией , HIF-1α и HIF-2α. В то время как HIF-1α стабилизирует p53, HIF-2α подавляет его. [ 33 ] Подавление р53 играет важную роль в фенотипе раковых стволовых клеток, индуцировании плюрипотентных стволовых клеток и других функциях и поведении стволовых клеток, таких как образование бластемы. Было показано, что клетки со сниженным уровнем р53 перепрограммируются в стволовые клетки с гораздо большей эффективностью, чем нормальные клетки. [ 34 ] [ 35 ] Исследования показывают, что отсутствие остановки клеточного цикла и апоптоза дает большему количеству клеток шанс перепрограммироваться. Также было показано, что снижение уровня р53 является решающим аспектом образования бластемы в ногах саламандр. [ 36 ] Регуляция р53 очень важна, действуя как барьер между стволовыми клетками и состоянием дифференцированных стволовых клеток, а также как барьер между функциональными и раковыми стволовыми клетками. [ 37 ]
Другой
[ редактировать ]
Помимо клеточных и молекулярных эффектов, описанных выше, р53 оказывает противораковое действие на тканевом уровне, которое действует путем ингибирования ангиогенеза . [ 38 ] По мере роста опухолей им необходимо рекрутировать новые кровеносные сосуды для их снабжения, и р53 ингибирует это, (i) вмешиваясь в работу регуляторов опухолевой гипоксии , которые также влияют на ангиогенез, таких как HIF1 и HIF2, (ii) ингибируя выработку факторов, способствующих ангиогенезу, и (iii) непосредственное увеличение продукции ингибиторов ангиогенеза, таких как аррестен . [ 39 ] [ 40 ]
p53, регулируя фактор ингибирования лейкемии, Было показано, что облегчает имплантацию мышам и, возможно, репродукцию человека. [ 41 ]
Иммунный ответ на инфекцию также включает р53 и NF-κB . Контроль контрольных точек клеточного цикла и апоптоза с помощью р53 ингибируется некоторыми инфекциями, такими как Mycoplasma , бактерии [ 42 ] повышая вероятность онкогенной инфекции .
Регулирование
[ редактировать ]p53 действует как сенсор клеточного стресса. Обычно его уровень поддерживается на низком уровне, поскольку он постоянно маркируется для деградации белком убиквитинлигазы E3 MDM2 . [ 43 ] p53 активируется в ответ на множество стрессоров, включая повреждение ДНК (вызванное УФ , ИК или химическими агентами, такими как перекись водорода), окислительный стресс , [ 44 ] осмотический шок , истощение рибонуклеотидов, вирусные инфекции легких [ 45 ] и дерегулированная экспрессия онкогенов. Эта активизация отмечена двумя важными событиями. Во-первых, период полураспада белка p53 резко увеличивается, что приводит к быстрому накоплению p53 в стрессированных клетках. Во-вторых, конформационные изменения заставляют р53 активироваться в качестве регулятора транскрипции в этих клетках. Критическим событием, приводящим к активации р53, является фосфорилирование его N-концевого домена. N-концевой домен активации транскрипции содержит большое количество сайтов фосфорилирования и может рассматриваться как основная мишень для протеинкиназ, передающих сигналы стресса.
Протеинкиназы , которые, как известно, нацелены на этот домен активации транскрипции р53, можно грубо разделить на две группы. Первая группа протеинкиназ принадлежит к семейству MAPK (JNK1-3, ERK1-2, p38 MAPK), которое, как известно, реагирует на несколько типов стресса, таких как повреждение мембран, окислительный стресс, осмотический шок, тепловой шок и т. д. Вторая группа протеинкиназ ( ATR , ATM , CHK1 и CHK2 , DNA-PK , CAK, TP53RK ) участвует в контрольной точке целостности генома, молекулярном каскаде, который обнаруживает и реагирует на несколько форм повреждений ДНК, вызванных генотоксическим стрессом. Онкогены также стимулируют активацию р53, опосредованную белком p14ARF .
В неиспытанных стрессом клетках уровень р53 поддерживается на низком уровне за счет непрерывной деградации р53. Белок под названием Mdm2 (также называемый HDM2 у людей) связывается с p53, предотвращая его действие, и транспортирует его из ядра в цитозоль . Mdm2 также действует как убиквитинлигаза и ковалентно присоединяет убиквитин к p53 и, таким образом, маркирует p53 для деградации протеасомой . Однако убиквитилирование р53 обратимо. При активации p53 также активируется Mdm2, настраивая контур обратной связи . Уровни p53 могут демонстрировать колебания (или повторяющиеся импульсы) в ответ на определенные стрессы, и эти импульсы могут иметь важное значение для определения того, переживут ли клетки стресс или погибнут. [ 46 ]
МИ-63 связывается с MDM2, реактивируя р53 в ситуациях, когда функция р53 подавляется. [ 47 ]
Убиквитин-специфическая протеаза, USP7 (или HAUSP ), может отщеплять убиквитин от p53, тем самым защищая его от протеасомно-зависимой деградации через путь убиквитинлигазы . Это один из способов стабилизации р53 в ответ на онкогенные повреждения. Также было показано, что USP42 деубиквитинирует р53 и может быть необходим для способности р53 реагировать на стресс. [ 48 ]
Недавние исследования показали, что HAUSP в основном локализуется в ядре, хотя некоторая его часть может быть обнаружена в цитоплазме и митохондриях. Сверхэкспрессия HAUSP приводит к стабилизации р53. Однако истощение HAUSP не приводит к снижению уровней p53, а скорее увеличивает уровни p53 из-за того, что HAUSP связывает и деубиквитинирует Mdm2. Было показано, что HAUSP является лучшим партнером по связыванию с Mdm2, чем p53, в нестрессированных клетках.
Однако было показано, что USP10 локализуется в цитоплазме нестрессовых клеток и деубиквитинирует цитоплазматический р53, обращая убиквитинирование Mdm2. После повреждения ДНК USP10 перемещается в ядро и способствует стабильности р53. Также USP10 не взаимодействует с Mdm2. [ 49 ]
Фосфорилирование N-конца р53 указанными выше протеинкиназами нарушает связывание Mdm2. Другие белки, такие как Pin1, затем рекрутируются на p53 и вызывают конформационные изменения p53, что еще больше предотвращает связывание Mdm2. Фосфорилирование также позволяет связывать коактиваторы транскрипции, такие как p300 и PCAF , которые затем ацетилируют C- конец p53, обнажая ДНК-связывающий домен p53, позволяя ему активировать или подавлять определенные гены. Ферменты деацетилазы, такие как Sirt1 и Sirt7 , могут деацетилировать р53, что приводит к ингибированию апоптоза. [ 50 ] Некоторые онкогены также могут стимулировать транскрипцию белков, связывающихся с MDM2, и ингибировать его активность.
Эпигенетические метки, такие как метилирование гистонов, также могут регулировать р53, например, р53 напрямую взаимодействует с репрессивным кофактором Trim24, который связывает гистоны в участках генома, которые эпигенетически репрессированы. [ 51 ] Trim24 предотвращает активацию p53 своих мишеней, но только в этих регионах, эффективно давая p53 возможность «считывать» профиль гистонов в ключевых генах-мишенях и действовать ген-специфичным образом.
Роль в болезни
[ редактировать ]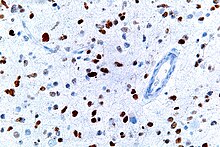
Если ген TP53 поврежден, подавление опухоли серьезно нарушено. У людей, которые наследуют только одну функциональную копию гена TP53 , скорее всего, в раннем взрослом возрасте разовьются опухоли — расстройство, известное как синдром Ли-Фраумени .
Ген TP53 также может быть модифицирован мутагенами ( химическими веществами , радиацией или вирусами ), что увеличивает вероятность неконтролируемого деления клеток. Более 50 процентов опухолей человека содержат мутацию или делецию гена TP53 . [ 52 ] Потеря р53 создает геномную нестабильность, которая чаще всего приводит к фенотипу анеуплоидии . [ 53 ]
Увеличение количества р53 может показаться решением для лечения опухолей или предотвращения их распространения. Однако это неприменимый метод лечения, поскольку он может вызвать преждевременное старение. [ 54 ] Восстановление эндогенной нормальной функции р53 имеет некоторые перспективы. Исследования показали, что это восстановление может привести к регрессии некоторых раковых клеток, не повреждая при этом другие клетки. Пути регрессии опухоли зависят главным образом от типа опухоли. Например, восстановление эндогенной функции р53 при лимфомах может вызвать апоптоз , в то время как рост клеток может снизиться до нормального уровня. Таким образом, фармакологическая реактивация р53 представляет собой жизнеспособный вариант лечения рака. [ 55 ] [ 56 ] Первая коммерческая генная терапия, Гендицин , была одобрена в Китае в 2003 году для лечения плоскоклеточного рака головы и шеи . Он доставляет функциональную копию гена p53 с помощью сконструированного аденовируса . [ 57 ]
Некоторые патогены также могут влиять на белок p53, который экспрессирует ген TP53 . Один из таких примеров, вирус папилломы человека (ВПЧ), кодирует белок E6, который связывается с белком p53 и инактивирует его. Этот механизм в сочетании с инактивацией регулятора клеточного цикла pRb белком Е7 ВПЧ обеспечивает повторное деление клеток, клинически проявляющееся в виде бородавок . Определенные типы ВПЧ, в частности типы 16 и 18, также могут приводить к прогрессированию доброкачественной бородавки к дисплазии шейки матки низкой или высокой степени , которые являются обратимыми формами предраковых поражений. Стойкая инфекция шейки матки на протяжении многих лет может вызвать необратимые изменения, ведущие к карциноме in situ и, в конечном итоге, к инвазивному раку шейки матки. Это является результатом воздействия генов ВПЧ, особенно тех, которые кодируют E6 и E7, которые представляют собой два вирусных онкопротеина, которые преимущественно сохраняются и экспрессируются при раке шейки матки путем интеграции вирусной ДНК в геном хозяина. [ 58 ]
Белок р53 постоянно вырабатывается и разлагается в клетках здоровых людей, что приводит к затуханию колебаний (см. стохастическую модель этого процесса в [ 59 ] ). Деградация белка р53 связана со связыванием MDM2. В петле отрицательной обратной связи сам MDM2 индуцируется белком p53. Мутантные белки p53 часто не способны индуцировать MDM2, что приводит к накоплению p53 на очень высоких уровнях. Более того, сам мутантный белок р53 может ингибировать нормальный уровень белка р53. Было показано, что в некоторых случаях одиночные миссенс-мутации в р53 нарушают стабильность и функцию р53. [ 60 ]
 На этом изображении показаны различные паттерны экспрессии р53 при раке эндометрия по данным хромогенной иммуногистохимии , из которых все, кроме дикого типа, по-разному называются аномальным/аберрантным/мутационным типом и являются надежными предикторами лежащей в основе мутации TP53: [ 61 ]
|

Показано, что подавление р53 в клетках рака молочной железы человека приводит к увеличению экспрессии гена хемокинового рецептора CXCR5 и активации миграции клеток в ответ на хемокин CXCL13 . [ 64 ]
Одно исследование показало, что белки p53 и Myc играют ключевую роль в выживании клеток хронического миелолейкоза (ХМЛ). Воздействие лекарств на белки p53 и Myc дало положительные результаты на мышах с ХМЛ. [ 65 ] [ 66 ]
Экспериментальный анализ мутаций р53
[ редактировать ]Большинство мутаций р53 выявляются с помощью секвенирования ДНК. Однако известно, что одиночные миссенс-мутации могут иметь широкий спектр функциональных эффектов от довольно легких до очень тяжелых. [ 60 ]
Широкий спектр фенотипов рака, обусловленных мутациями гена TP53 , также подтверждается тем фактом, что разные изоформы белков p53 имеют разные клеточные механизмы профилактики рака. Мутации в TP53 могут привести к появлению различных изоформ, предотвращая их общую функциональность в различных клеточных механизмах и тем самым расширяя фенотип рака от легкого до тяжелого. Недавние исследования показывают, что изоформы р53 по-разному экспрессируются в разных тканях человека, а мутации в отношении потери или усиления функции внутри изоформ могут вызывать тканеспецифичный рак или обеспечивать раковых стволовых клеток потенциал в различных тканях. [ 11 ] [ 67 ] [ 68 ] [ 69 ] Мутация TP53 также влияет на энергетический обмен и увеличивает гликолиз в клетках рака молочной железы. [ 70 ]
Динамика белков p53 вместе с его антагонистом Mdm2 указывает на то, что уровни p53 в единицах концентрации колеблются в зависимости от времени. Это « затухающее » колебание клинически задокументировано. [ 71 ] и математически смоделированы . [ 72 ] [ 73 ] Математические модели также показывают, что концентрация p53 колеблется намного быстрее, когда тератогены, такие как двухцепочечные разрывы (DSB) или УФ-излучение вводятся в систему . Это подтверждает и моделирует нынешнее понимание динамики р53, когда повреждение ДНК вызывает активацию р53 ( см. в разделе «Регуляция р53» дополнительную информацию ). Современные модели также могут быть полезны для моделирования мутаций изоформ р53 и их влияния на колебания р53, тем самым способствуя препаратов de novo тканеспецифичных фармакологических открытию .
Открытие
[ редактировать ]p53 был идентифицирован в 1979 году Лайонелом Кроуфордом , Дэвидом П. Лейном , Арнольдом Левином и Ллойдом Олдом , работавшими в Имперском фонде исследования рака (Великобритания), Принстонском университете /UMDNJ (Институт рака Нью-Джерси) и Мемориальном онкологическом центре Слоана-Кеттеринга соответственно. . Ранее предполагалось, что он существует как мишень вируса SV40 , штамма, вызывающего развитие опухолей. Название p53 было дано в 1979 году для описания кажущейся молекулярной массы .
Ген TP53 мыши был впервые клонирован Петром Чумаковым из Академии наук СССР в 1982 году. [ 74 ] и независимо в 1983 году Моше Ореном в сотрудничестве с Дэвидом Гиволом ( Институт науки Вейцмана ). [ 75 ] [ 76 ] человека Ген TP53 был клонирован в 1984 году. [ 7 ] и полноформатный клон в 1985 году. [ 77 ]
Первоначально предполагалось, что он является онкогеном из-за использования мутированной кДНК после очистки мРНК опухолевых клеток . Его роль как гена-супрессора опухолей была раскрыта в 1989 году Бертом Фогельштейном из Медицинской школы Джонса Хопкинса и Арнольдом Левином из Принстонского университета. [ 78 ] [ 79 ] p53 был впоследствии идентифицирован как фактор транскрипции Гильерминой Лозано, работающей в Онкологическом центре доктора медицины Андерсона . [ 80 ]
Уоррен Мальцман из Института Ваксмана Университета Рутгерса впервые продемонстрировал, что TP53 реагирует на повреждение ДНК в виде ультрафиолетового излучения. [ 81 ] В серии публикаций 1991–92 годов Майкл Кастан из Университета Джонса Хопкинса сообщил, что TP53 является важной частью пути передачи сигнала, который помогает клеткам реагировать на повреждение ДНК. [ 82 ]
назвал р53 молекулой года В 1993 году журнал Science . [ 83 ]
Структура
[ редактировать ]

p53 имеет семь доменов :
- кислый N-концевой домен активации транскрипции (TAD), также известный как домен активации 1 (AD1), который активирует факторы транскрипции . N-конец содержит два комплементарных домена активации транскрипции, главный из которых находится в остатках 1–42, а второстепенный — в остатках 55–75, специфически участвующих в регуляции нескольких проапоптотических генов. [ 84 ]
- домен активации 2 (AD2), важный для апоптотической активности: остатки 43–63.
- Богатый пролином домен, важный для апоптотической активности р53 путем ядерного экспорта через МАРК : остатки 64–92.
- центральный ДНК -связывающий коровый домен ( DBD ). Содержит один атом цинка и несколько аргинина аминокислот : остатки 102–292. Эта область отвечает за связывание ко-репрессора р53 LMO3 . [ 85 ]
- Домен передачи сигналов ядерной локализации (NLS), остатки 316–325.
- домен гомоолигомеризации (OD): остатки 307–355. Тетрамеризация необходима для активности р53 in vivo .
- С-конец участвует в подавлении связывания ДНК центрального домена: остатки 356–393. [ 86 ]
Мутации, деактивирующие р53 при раке, обычно возникают в DBD. Большинство этих мутаций разрушают способность белка связываться с целевыми последовательностями ДНК и, таким образом, предотвращают транскрипционную активацию этих генов. Таким образом, мутации в DBD представляют собой рецессивные мутации с потерей функции . Молекулы р53 с мутациями в OD димеризуются с р53 дикого типа и не позволяют им активировать транскрипцию. Следовательно, мутации OD оказывают доминирующее негативное влияние на функцию р53.
p53 дикого типа представляет собой лабильный белок , содержащий свернутые и неструктурированные области , которые действуют синергично. [ 87 ]
Анализ SDS-PAGE показывает, что p53 представляет собой белок массой 53 килодальтон (кДа). Однако фактическая масса полноразмерного белка р53 (p53α), рассчитанная по сумме масс аминокислотных остатков, составляет всего 43,7 кДа. Эта разница обусловлена большим количеством остатков пролина в белке, которые замедляют его миграцию при SDS-PAGE, в результате чего он кажется тяжелее, чем есть на самом деле. [ 88 ]
Изоформы
[ редактировать ]Как и 95% генов человека, TP53 кодирует более одного белка. Все эти белки р53 называются изоформами р53 . [ 5 ] Размер этих белков варьируется от 3,5 до 43,7 кДа. Несколько изоформ были открыты в 2005 году, и на данный момент идентифицировано 12 изоформ р53 человека (p53α, p53β, p53γ, ∆40p53α, ∆40p53β, ∆40p53γ, ∆133p53α, ∆133p53β, ∆133p53γ, ∆160p53α, ∆160p53β, ∆160p53γ). Более того, изоформы р53 экспрессируются тканезависимым образом, а р53α никогда не экспрессируется отдельно. [ 11 ]
Полноразмерные белки изоформы р53 можно разделить на различные белковые домены . Начиная с N-конца , сначала идут аминоконцевые домены активации транскрипции (TAD 1, TAD 2), которые необходимы для индукции подмножества генов-мишеней p53. За этим доменом следует домен, богатый пролином (PXXP), в результате чего мотив PXXP повторяется (P представляет собой пролин, а X может быть любой аминокислотой). Он необходим, среди прочего, для апоптоза, опосредованного р53 . [ 89 ] У некоторых изоформ отсутствует богатый пролином домен, например Δ133p53β,γ и Δ160p53α,β,γ; следовательно, некоторые изоформы р53 не опосредуют апоптоз, что подчеркивает разнообразную роль гена TP53 . [ 67 ] Далее следует ДНК-связывающий домен (DBD), который позволяет белкам секвенировать специфическое связывание. домен С-концевой завершает белок. Он включает сигнал ядерной локализации (NLS), сигнал ядерного экспорта (NES) и домен олигомеризации (OD). NLS и NES отвечают за субклеточную регуляцию р53. Через OD р53 может образовывать тетрамер, а затем связываться с ДНК. Среди изоформ некоторые домены могут отсутствовать, но все они имеют большую часть высококонсервативного ДНК-связывающего домена.
Изоформы образуются по разным механизмам. Бета- и гамма-изоформы образуются в результате множественного сплайсинга интрона 9, что приводит к образованию другого С-конца. Кроме того, использование внутреннего промотора в интроне 4 приводит к появлению изоформ ∆133 и ∆160, в которых отсутствует домен TAD и часть DBD. Более того, альтернативная инициация трансляции на кодоне 40 или 160 несет изоформы ∆40p53 и ∆160p53. [ 11 ]
Из-за изоформной природы белков р53 было несколько источников доказательств, показывающих, что мутации в гене TP53 , приводящие к возникновению мутированных изоформ, являются возбудителями различных фенотипов рака, от легких до тяжелых, из-за одиночной мутации в гене TP53 ( более подробную информацию см. в разделе «Экспериментальный анализ мутаций р53» ).
Взаимодействия
[ редактировать ]Было показано, что p53 взаимодействует с:
- АИМП2 , [ 90 ]
- АНКРД2 , [ 91 ]
- АПТХ , [ 92 ]
- банкомат , [ 93 ] [ 94 ] [ 95 ] [ 96 ] [ 97 ]
- АТР , [ 93 ] [ 94 ]
- АТФ3 , [ 98 ] [ 99 ]
- AURKAПРОТИВ [ 100 ]
- БАК1 , [ 101 ]
- БАРД1 , [ 102 ]
- БЛМ , [ 103 ] [ 104 ] [ 105 ] [ 106 ]
- БРЦА1 , [ 102 ] [ 107 ] [ 108 ] [ 109 ] [ 110 ]
- БРЦА2 , [ 102 ] [ 111 ]
- БРЦЦ3 , [ 102 ]
- БРЭ , [ 102 ]
- ЦЭБПЗ , [ 112 ]
- CDC14A , [ 113 ]
- CDK1 , [ 114 ] [ 115 ]
- ЦФЛАР , [ 116 ]
- ЧЕК1 , [ 103 ] [ 117 ] [ 118 ]
- CCNG1 , [ 119 ]
- КРЕББП , [ 120 ] [ 121 ] [ 122 ]
- CREB1 , [ 122 ]
- Циклин Х , [ 123 ]
- СДК7 , [ 123 ] [ 124 ]
- ДНК-ПКкс , [ 94 ] [ 117 ] [ 125 ]
- Е4F1 , [ 126 ] [ 127 ]
- ЭФЕМП2 , [ 128 ]
- ЭИФ2АК2 , [ 129 ]
- ОН , [ 130 ]
- ЭП300 , [ 121 ] [ 131 ] [ 132 ] [ 133 ]
- ЭРСС6 , [ 134 ] [ 135 ]
- ГНЛ3 , [ 136 ]
- GPS2 , [ 137 ]
- ГСК3Б , [ 138 ]
- ХСП90АА1 , [ 139 ] [ 140 ] [ 141 ]
- ХИФ1А , [ 142 ] [ 143 ] [ 144 ] [ 145 ]
- ХИПК1 , [ 146 ]
- ХИПК2 , [ 147 ] [ 148 ]
- ХМГБ1 , [ 149 ] [ 150 ]
- HSPA9 , [ 151 ]
- Хантингтин , [ 152 ]
- ИНГ1 , [ 153 ] [ 154 ]
- ИНГ4 , [ 155 ] [ 156 ]
- ИНГ5 , [ 155 ]
- IκBα , [ 157 ]
- КПНБ1 , [ 139 ]
- ЛМО3 , [ 85 ]
- Мдм2 , [ 120 ] [ 158 ] [ 159 ] [ 160 ]
- МДМ4 , [ 161 ] [ 162 ]
- МЕД1 , [ 163 ] [ 164 ]
- МАПК9 , [ 165 ] [ 166 ]
- МНАТ1 , [ 124 ]
- НДН , [ 167 ]
- НКЛ , [ 168 ]
- ОНЕМЕВШИЙ , [ 169 ]
- НФ-кБ , [ 170 ]
- П16 , [ 126 ] [ 160 ] [ 171 ]
- ПАРК , [ 172 ]
- ПАРП1 , [ 92 ] [ 173 ]
- ПИАС1 , [ 128 ] [ 174 ]
- CDC14B , [ 113 ]
- ПИН1 , [ 175 ] [ 176 ]
- ПЛАГЛ1 , [ 177 ]
- ПЛК3 , [ 178 ] [ 179 ]
- ПРКРА , [ 180 ]
- ПОБ , [ 181 ]
- ПМЛ , [ 158 ] [ 182 ] [ 183 ]
- ПСМЕ3 , [ 184 ]
- ПТЭН , [ 159 ]
- ПТК2 , [ 185 ]
- ПТТГ1 , [ 186 ]
- РАД51 , [ 102 ] [ 187 ] [ 188 ]
- РЧИ1 , [ 189 ] [ 190 ]
- РЕЛА , [ 170 ]
- Я подавляю [ нужна ссылка ]
- РПА1 , [ 191 ] [ 192 ]
- РПЛ11 , [ 171 ]
- С100Б , [ 193 ]
- СУМО1 , [ 194 ] [ 195 ]
- СМАРКА4 , [ 196 ]
- SMARCB1 , [ 196 ]
- СМН1 , [ 197 ]
- СТАТ3 , [ 170 ]
- ТБП , [ 198 ] [ 199 ]
- ТФАП2А , [ 200 ]
- ТФДП1 , [ 201 ]
- ТИГР , [ 202 ]
- ТОП1 , [ 203 ] [ 204 ]
- ТОП2А , [ 205 ]
- ТП53БП1 , [ 103 ] [ 206 ] [ 207 ] [ 208 ] [ 209 ] [ 210 ] [ 211 ]
- ТП53БП2 , [ 211 ] [ 212 ]
- ТОП2Б , [ 205 ]
- ТП53ИНП1 , [ 213 ] [ 214 ]
- ТСГ101 , [ 215 ]
- БЕ2А , [ 216 ]
- БЭ2И , [ 128 ] [ 194 ] [ 217 ] [ 218 ]
- ЮБК , [ 90 ] [ 184 ] [ 195 ] [ 219 ] [ 220 ] [ 221 ] [ 222 ] [ 223 ]
- УСП7 , [ 224 ]
- USP10 , [ 49 ]
- ВРН , [ 106 ] [ 225 ]
- WWOX , [ 226 ]
- XPB , [ 134 ]
- YBX1 , [ 91 ] [ 227 ]
- ЮПЕЛ3 , [ 228 ]
- ЯВАЗ , [ 229 ]
- Зиф268 , [ 230 ]
- ЗНФ148 , [ 231 ]
- СИРТ1 , [ 232 ]
- циркРНК_014511. [ 233 ]
См. также
[ редактировать ]- Пифитрин , ингибитор Р53.
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000141510 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000059552 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с Суржет С., член парламента Хури, Дж. К. Бурдон (декабрь 2013 г.). «Раскрытие роли вариантов сплайсинга р53 в злокачественных новообразованиях человека: клиническая перспектива» . Онкомишени и терапия . 7 : 57–68. дои : 10.2147/OTT.S53876 . ПМЦ 3872270 . ПМИД 24379683 .
- ^ Туфекчан Э, Толедо Ф (май 2018 г.). «Возвращение к «Хранителю генома»: p53 подавляет активность генов, необходимых для поддержания теломер, восстановления ДНК и структуры центромер» . Раки . 10 (5): 135. doi : 10.3390/cancers10050135 . ПМЦ 5977108 . ПМИД 29734785 .
- ^ Jump up to: а б с Матлашевски Г., Лэмб П., Пим Д. и др. (декабрь 1984 г.). «Выделение и характеристика клона кДНК р53 человека: экспрессия гена р53 человека» . Журнал ЭМБО . 3 (13): 3257–62. дои : 10.1002/j.1460-2075.1984.tb02287.x . ПМЦ 557846 . ПМИД 6396087 .
- ^ Jump up to: а б Изобе М., Эмануэль Б.С., Гивол Д. и др. (1986). «Локализация гена опухолевого антигена p53 человека на полосе 17p13». Природа . 320 (6057): 84–5. Бибкод : 1986Natur.320...84I . дои : 10.1038/320084a0 . ПМИД 3456488 . S2CID 4310476 .
- ^ Jump up to: а б Керн С.Е., Кинцлер К.В., Брускин А. и др. (июнь 1991 г.). «Идентификация p53 как ДНК-связывающего белка, специфичного для последовательности». Наука . 252 (5013): 1708–11. Бибкод : 1991Sci...252.1708K . дои : 10.1126/science.2047879 . ПМИД 2047879 . S2CID 19647885 .
- ^ Jump up to: а б Макбрайд О.В., Мерри Д., Гивол Д. (январь 1986 г.). «Ген клеточного опухолевого антигена p53 человека расположен на коротком плече 17 хромосомы (17p13)» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (1): 130–4. Бибкод : 1986ПНАС...83..130М . дои : 10.1073/pnas.83.1.130 . ПМК 322805 . ПМИД 3001719 .
- ^ Jump up to: а б с д Бурдон Дж.К., Фернандес К., Мюррей-Змиевски Ф. и др. (сентябрь 2005 г.). «Изоформы р53 могут регулировать транскрипционную активность р53» . Гены и развитие . 19 (18): 2122–37. дои : 10.1101/gad.1339905 . ПМЦ 1221884 . ПМИД 16131611 .
- ^ Левин А.Дж., Лейн Д.П., ред. (2010). Семейство р53 . Перспективы Колд-Спринг-Харбор в биологии. Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-830-0 .
- ^ Член парламента Хури, Бурдон Дж.К. Изоформы p53: внутриклеточный микропроцессор? Гены рака. 2011, апрель 2(4): 453-65. дои: 10.1177/1947601911408893. PMID: 21779513; PMCID: PMC3135639.
- ^ Мэй П, Мэй Е (декабрь 1999 г.). «Двадцать лет исследований р53: структурные и функциональные аспекты белка р53» . Онкоген . 18 (53): 7621–36. дои : 10.1038/sj.onc.1203285 . ПМИД 10618702 .
- ^ «Филогенетический маркер OrthoMaM: кодирующая последовательность TP53» . Архивировано из оригинала 17 марта 2018 г. Проверено 2 декабря 2009 г.
- ^ Клуг С.Дж., Рессинг М., Кениг Дж. и др. (август 2009 г.). «Полиморфизм кодона 72 TP53 и рак шейки матки: объединенный анализ отдельных данных из 49 исследований». «Ланцет». Онкология . 10 (8): 772–84. дои : 10.1016/S1470-2045(09)70187-1 . ПМИД 19625214 .
- ^ Сонояма Т., Сакаи А., Мита Ю. и др. (2011). «Полиморфизм кодона 72 TP53 связан с риском рака поджелудочной железы у мужчин, курильщиков и пьющих» . Отчеты о молекулярной медицине . 4 (3): 489–95. дои : 10.3892/ммр.2011.449 . ПМИД 21468597 .
- ^ Алавади С., Габро Л., Алсалех М. и др. (сентябрь 2011 г.). «Полиморфизм гена P53 и риск рака молочной железы у арабских женщин». Медицинская онкология . 28 (3): 709–15. дои : 10.1007/s12032-010-9505-4 . ПМИД 20443084 . S2CID 207372095 .
- ^ Ю Х., Хуан Ю.Дж., Лю Цз. и др. (сентябрь 2011 г.). «Влияние полиморфизма промотора MDM2 и полиморфизма кодона 72 p53 на риск и возраст начала плоскоклеточного рака головы и шеи» . Молекулярный канцерогенез . 50 (9): 697–706. дои : 10.1002/mc.20806 . ПМЦ 3142329 . ПМИД 21656578 .
- ^ Пяо Дж.М., Ким Х.Н., Сонг Х.Р. и др. (сентябрь 2011 г.). «Полиморфизм кодона 72 p53 и риск рака легких у корейского населения». Рак легких . 73 (3): 264–7. дои : 10.1016/j.lungcan.2010.12.017 . ПМИД 21316118 .
- ^ Ван Дж.Дж., Чжэн Ю., Сунь Л. и др. (ноябрь 2011 г.). «Полиморфизм кодона 72 TP53 и предрасположенность к колоректальному раку: метаанализ». Отчеты по молекулярной биологии . 38 (8): 4847–53. дои : 10.1007/s11033-010-0619-8 . ПМИД 21140221 . S2CID 11730631 .
- ^ Цзян Д.К., Яо Л., Рен В.Х. и др. (декабрь 2011 г.). «Полиморфизм TP53 Arg72Pro и риск рака эндометрия: метаанализ». Медицинская онкология . 28 (4): 1129–35. дои : 10.1007/s12032-010-9597-x . ПМИД 20552298 . S2CID 32990396 .
- ^ Туроу Х.С., Хаак Р., Хартвиг Ф.П. и др. (декабрь 2011 г.). «Полиморфизм гена TP53: значение для рака, этнической принадлежности и веса при рождении в бразильской когорте». Журнал биологических наук . 36 (5): 823–31. дои : 10.1007/s12038-011-9147-5 . ПМИД 22116280 . S2CID 23027087 .
- ^ Хуан С.И., Су К.Т., Чу Дж.С. и др. (декабрь 2011 г.). «Полиморфизмы кодона 72 P53 и SNP309 MDM2 и риск почечно-клеточного рака в зоне с низким уровнем воздействия мышьяка». Токсикология и прикладная фармакология . 257 (3): 349–55. Бибкод : 2011ToxAP.257..349H . дои : 10.1016/j.taap.2011.09.018 . ПМИД 21982800 .
- ^ Гилберт С.Ф. Биология развития, 10-е изд . Сандерленд, Массачусетс, США: Издательство Sinauer Associates, Inc. п. 588.
- ^ Национальный центр биотехнологической информации (1998). «Кожа и соединительная ткань» . Гены и болезни . Национальные институты здравоохранения США . Проверено 28 мая 2008 г.
- ^ Бейтс С., Филлипс А.С., Кларк П.А. и др. (сентябрь 1998 г.). «p14ARF связывает опухолевые супрессоры RB и p53». Природа . 395 (6698): 124–5. Бибкод : 1998Natur.395..124B . дои : 10.1038/25867 . ПМИД 9744267 . S2CID 4355786 .
- ^ «Хранитель Генома начал загорать» . Новый учёный . 17 марта 2007 года . Проверено 29 марта 2007 г.
- ^ Куи Р., Видлунд Х.Р., Файги Э. и др. (март 2007 г.). «Центральная роль р53 в реакции на загар и патологической гиперпигментации» . Клетка . 128 (5): 853–64. дои : 10.1016/j.cell.2006.12.045 . ПМИД 17350573 .
- ^ Jump up to: а б Джайн А.К., Олтон К., Яковино М. и др. (2012). «p53 регулирует клеточный цикл и микроРНК, способствуя дифференцировке эмбриональных стволовых клеток человека» . ПЛОС Биология . 10 (2): e1001268. дои : 10.1371/journal.pbio.1001268 . ПМК 3289600 . ПМИД 22389628 .
- ^ Майметс Т., Неганова И., Армстронг Л. и др. (сентябрь 2008 г.). «Активация р53 нутлином приводит к быстрой дифференцировке эмбриональных стволовых клеток человека» . Онкоген . 27 (40): 5277–87. дои : 10.1038/onc.2008.166 . ПМИД 18521083 .
- ^ тер Хуурне М., Пэн Т., Йи Г. и др. (февраль 2020 г.). «Критическая роль P53 в регуляции клеточного цикла эмбриональных стволовых клеток основного состояния» . Отчеты о стволовых клетках . 14 (2): 175–183. doi : 10.1016/j.stemcr.2020.01.001 . ПМК 7013234 . ПМИД 32004494 .
- ^ Дас Б., Баят-Мохтари Р., Цуй М. и др. (август 2012 г.). «HIF-2α подавляет р53, повышая стволовость и регенеративный потенциал эмбриональных стволовых клеток человека» . Стволовые клетки . 30 (8): 1685–95. дои : 10.1002/stem.1142 . ПМЦ 3584519 . ПМИД 22689594 .
- ^ Лейк Б.Б., Финк Дж., Клеметсауне Л. и др. (май 2012 г.). «Контекст-зависимое усиление индуцированного перепрограммирования плюрипотентных стволовых клеток путем подавления Puma» . Стволовые клетки . 30 (5): 888–97. дои : 10.1002/stem.1054 . ПМК 3531606 . ПМИД 22311782 .
- ^ Марион Р.М., Страти К., Ли Х. и др. (август 2009 г.). «Реакция на повреждение ДНК, опосредованная р53, ограничивает перепрограммирование для обеспечения геномной целостности iPS-клеток» . Природа . 460 (7259): 1149–53. Бибкод : 2009Natur.460.1149M . дои : 10.1038/nature08287 . ПМК 3624089 . ПМИД 19668189 .
- ^ Юн М.Х., Гейтс П.Б., Брокс Дж.П. (октябрь 2013 г.). «Регуляция р53 имеет решающее значение для регенерации конечностей позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (43): 17392–7. Бибкод : 2013PNAS..11017392Y . дои : 10.1073/pnas.1310519110 . ПМЦ 3808590 . ПМИД 24101460 .
- ^ Алони-Гринштейн Р., Шетцер Ю., Кауфман Т. и др. (август 2014 г.). «p53: барьер на пути образования раковых стволовых клеток» . Письма ФЭБС . 588 (16): 2580–9. Бибкод : 2014FEBSL.588.2580A . дои : 10.1016/j.febslet.2014.02.011 . ПМИД 24560790 . S2CID 37901173 .
- ^ Jump up to: а б Бабаи Г., Алиараб А., Асгари Востоколаи М. и др. (ноябрь 2021 г.). «Перекрестная связь между p53 и метастазами: фокус на эпителиально-мезенхимальном переходе, раковых стволовых клетках, ангиогенезе, аутофагии и аноикисе». Отчеты по молекулярной биологии . 48 (11): 7545–7557. дои : 10.1007/s11033-021-06706-1 . ПМИД 34519942 . S2CID 237506513 .
- ^ Теодоро Дж.Г., Эванс С.К., Грин М.Р. (ноябрь 2007 г.). «Ингибирование опухолевого ангиогенеза с помощью р53: новая роль хранителя генома». Журнал молекулярной медицины (обзор). 85 (11): 1175–1186. дои : 10.1007/s00109-007-0221-2 . ПМИД 17589818 . S2CID 10094554 .
- ^ Асадиан С., Эль-Ассаад В., Ван XQ и др. (март 2012 г.). «p53 ингибирует ангиогенез, индуцируя выработку аррестена» . Исследования рака . 72 (5): 1270–1279. дои : 10.1158/0008-5472.CAN-11-2348 . ПМИД 22253229 .
- ^ Ху В., Фэн З., Терески А.К. и др. (ноябрь 2007 г.). «p53 регулирует материнское воспроизводство посредством LIF». Природа . 450 (7170): 721–4. Бибкод : 2007Natur.450..721H . дои : 10.1038/nature05993 . ПМИД 18046411 . S2CID 4357527 .
- ^ Борхсениус С.Н., Дакс А., Федорова О. и др. (январь 2018 г.). «Влияние микоплазменной инфекции на реакцию организма хозяина посредством передачи сигналов p53/NF-κB». Журнал клеточной физиологии . 234 (1): 171–180. дои : 10.1002/jcp.26781 . ПМИД 30146800 .
- ^ Быков В.Я., Эрикссон С.Е., Бьянки Дж. и др. (февраль 2018 г.). «Нацеливание на мутантный р53 для эффективной терапии рака». Обзоры природы. Рак . 18 (2): 89–102. дои : 10.1038/nrc.2017.109 . ПМИД 29242642 . S2CID 4552678 .
- ^ Хан Э.С., Мюллер Ф.Л., Перес VI и др. (июнь 2008 г.). «Признак экспрессии генов in vivo, вызывающий окислительный стресс» . Физиологическая геномика . 34 (1): 112–126. doi : 10.1152/физиологгеномика.00239.2007 . ПМЦ 2532791 . ПМИД 18445702 .
- ^ Грахалес-Рейес Г.Е., Колонна М. (август 2020 г.). «Интерфероновые реакции при вирусных пневмониях». Наука . 369 (6504): 626–627. Бибкод : 2020Sci...369..626G . дои : 10.1126/science.abd2208 . ПМИД 32764056 .
- ^ Первис Дж.Э., Кархос К.В., Мок С. и др. (июнь 2012 г.). «Динамика p53 контролирует судьбу клеток» . Наука . 336 (6087): 1440–1444. Бибкод : 2012Sci...336.1440P . дои : 10.1126/science.1218351 . ПМК 4162876 . ПМИД 22700930 .
- ^ Каннер Дж.А., Собо М., Болл С. и др. (сентябрь 2009 г.). «МИ-63: новый низкомолекулярный ингибитор нацелен на MDM2 и индуцирует апоптоз в эмбриональных и альвеолярных клетках рабдомиосаркомы с р53 дикого типа» . Британский журнал рака . 101 (5): 774–81. дои : 10.1038/sj.bjc.6605199 . ПМЦ 2736841 . ПМИД 19707204 .
- ^ Хок А.К., Виньерон А.М., Картер С. и др. (ноябрь 2011 г.). «Регуляция стабильности и функции р53 с помощью деубиквитинирующего фермента USP42» . Журнал ЭМБО . 30 (24): 4921–30. дои : 10.1038/emboj.2011.419 . ПМЦ 3243628 . ПМИД 22085928 .
- ^ Jump up to: а б Юань Дж., Луо К., Чжан Л. и др. (февраль 2010 г.). «USP10 регулирует локализацию и стабильность p53 путем деубиквитинирования p53» . Клетка . 140 (3): 384–396. дои : 10.1016/j.cell.2009.12.032 . ПМК 2820153 . ПМИД 20096447 .
- ^ Вахрушева О., Смолка С., Гаджавада П. и др. (март 2008 г.). «Sirt7 повышает стрессоустойчивость кардиомиоцитов и предотвращает апоптоз и воспалительную кардиомиопатию у мышей» . Исследование кровообращения . 102 (6): 703–10. дои : 10.1161/CIRCRESAHA.107.164558 . ПМИД 18239138 .
- ^ Исбель Л., Искар М., Дурду С. и др. (июнь 2023 г.). «Считывание метилирования гистонов с помощью Trim24 локально ограничивает открытие хроматина с помощью p53» . Структурная и молекулярная биология природы . 30 (7): 948–57. дои : 10.1038/s41594-023-01021-8 . hdl : 2440/139184 . ПМЦ 10352137 . ПМИД 37386214 .
- ^ Хольштейн М., Сидранский Д., Фогельштейн Б. и др. (июль 1991 г.). «Мутации р53 при раке человека» . Наука . 253 (5015): 49–53. Бибкод : 1991Sci...253...49H . дои : 10.1126/science.1905840 . ПМИД 1905840 . S2CID 38527914 .
- ^ Шмитт К.А., Фридман Дж.С., Ян М. и др. (апрель 2002 г.). «Раскрытие функций супрессора опухоли p53 in vivo» . Раковая клетка . 1 (3): 289–98. дои : 10.1016/S1535-6108(02)00047-8 . ПМИД 12086865 .
- ^ Тайнер С.Д., Венкатачалам С., Чой Дж. и др. (январь 2002 г.). «Мутантные мыши p53, демонстрирующие фенотипы, связанные с ранним старением». Природа . 415 (6867): 45–53. Бибкод : 2002Natur.415...45T . дои : 10.1038/415045а . ПМИД 11780111 . S2CID 749047 .
- ^ Вентура А., Кирш Д.Г., Маклафлин М.Е. и др. (февраль 2007 г.). «Восстановление функции р53 приводит к регрессии опухоли in vivo». Природа . 445 (7128): 661–5. дои : 10.1038/nature05541 . ПМИД 17251932 . S2CID 4373520 .
- ^ Герце Х.Д., Денг В., Хельма Дж. и др. (2013). «Визуализация и целенаправленное нарушение белковых взаимодействий в живых клетках» . Природные коммуникации . 4 : 2660. Бибкод : 2013NatCo...4.2660H . дои : 10.1038/ncomms3660 . ПМЦ 3826628 . ПМИД 24154492 .
- ^ Пирсон С., Цзя Х., Кандачи К. (январь 2004 г.). «Китай одобряет первую генную терапию» . Природная биотехнология . 22 (1): 3–4. дои : 10.1038/nbt0104-3 . ПМК 7097065 . ПМИД 14704685 .
- ^ Анджелетти ПК, Чжан Л., Вуд С. (2008). «Вирусная этиология злокачественных опухолей, связанных со СПИДом». ВИЧ-1: молекулярная биология и патогенез . Достижения фармакологии. Том. 56. стр. 509–57. дои : 10.1016/S1054-3589(07)56016-3 . ISBN 978-0-12-373601-7 . ПМК 2149907 . ПМИД 18086422 .
- ^ Рибейро А.С., Шарлебуа Д.А., Ллойд-Прайс Дж. (декабрь 2007 г.). «CellLine, симулятор стохастического клеточного происхождения» . Биоинформатика . 23 (24): 3409–3411. doi : 10.1093/биоинформатика/btm491 . ПМИД 17928303 .
- ^ Jump up to: а б Буллок А.Н., Хенкель Дж., ДеДекер Б.С. и др. (декабрь 1997 г.). «Термодинамическая стабильность корового домена р53 дикого типа и мутанта» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (26): 14338–42. Бибкод : 1997PNAS...9414338B . дои : 10.1073/pnas.94.26.14338 . ПМК 24967 . ПМИД 9405613 .
- ^ Кёбель М., Роннетт Б.М., Сингх Н. и др. (январь 2019 г.). «Интерпретация иммуногистохимии P53 при карциноме эндометрия: на пути к повышению воспроизводимости» . Международный журнал гинекологической патологии . 38 (Приложение 1): S123–S131. дои : 10.1097/PGP.0000000000000488 . ПМК 6127005 . ПМИД 29517499 .
 В эту статью включен текст, доступный по лицензии CC BY 4.0 .
В эту статью включен текст, доступный по лицензии CC BY 4.0 .
- ^ Изображение взято из следующего источника с некоторыми изменениями Микаэля Хэггстрема, доктора медицинских наук:
- Шалленберг С., Плаж Х., Хофбауэр С. и др. (2023). «Измененная экспрессия p53/p16 связана с прогрессированием уротелиальной карциномы, но в значительной степени не связана с прогнозом при мышечно-инвазивных опухолях» . Акта Онкол . 62 (12): 1880–1889. дои : 10.1080/0284186X.2023.2277344 . ПМИД 37938166 . - ^ Источник роли в различении PUNLMP от рака низкой степени злокачественности:
- Калантари М.Р., Ахмадния Х (2007). «Сверхэкспрессия P53 при уротелиальных новообразованиях мочевого пузыря: новый аспект классификации Всемирной организации здравоохранения / Международного общества урологической патологии» . Урол Дж . 4 (4): 230–3. ПМИД 18270948 . - ^ Миткин Н.А., Хук К.Д., Шварц А.М. и др. (март 2015 г.). «p53-зависимая экспрессия хемокинового рецептора CXCR5 в клетках рака молочной железы MCF-7» . Научные отчеты . 5 (5): 9330. Бибкод : 2015NatSR...5E9330M . дои : 10.1038/srep09330 . ПМК 4365401 . ПМИД 25786345 .
- ^ Авраам С.А., Хопкрофт Л.Е., Каррик Э. и др. (июнь 2016 г.). «Двойное воздействие на p53 и c-MYC избирательно уничтожает лейкозные стволовые клетки» . Природа . 534 (7607): 341–6. Бибкод : 2016Natur.534..341A . дои : 10.1038/nature18288 . ПМЦ 4913876 . ПМИД 27281222 .
- ^ «Ученые идентифицируют лекарства, воздействующие на «ахиллесову пяту» клеток хронического миелолейкоза» . мояНаука . 08.06.2016 . Проверено 9 июня 2016 г.
- ^ Jump up to: а б Член парламента Хури, Дж. К. Бурдон (апрель 2011 г.). «Изоформы p53: внутриклеточный микропроцессор?» . Гены и рак . 2 (4): 453–65. дои : 10.1177/1947601911408893 . ПМЦ 3135639 . ПМИД 21779513 .
- ^ Эйвери-Кейда К.А., Мортен Б., Вонг-Браун М.В. и др. (март 2014 г.). «Относительная экспрессия мРНК изоформ р53 при раке молочной железы связана с клиническими особенностями и исходом» . Канцерогенез . 35 (3): 586–96. дои : 10.1093/carcin/bgt411 . ПМИД 24336193 .
- ^ Арсик Н., Гадеа Г., Лагерквист Э.Л. и др. (апрель 2015 г.). «Изоформа p53 Δ133p53β повышает потенциал раковых стволовых клеток» . Отчеты о стволовых клетках . 4 (4): 531–40. дои : 10.1016/j.stemcr.2015.02.001 . ПМК 4400643 . ПМИД 25754205 .
- ^ Харами-Папп Х., Понгор Л.С., Мункачи Г. и др. (октябрь 2016 г.). «Мутация TP53 влияет на энергетический обмен и увеличивает гликолиз при раке молочной железы» . Онкотаргет . 7 (41): 67183–67195. дои : 10.18632/oncotarget.11594 . ПМК 5341867 . ПМИД 27582538 .
- ^ Гева-Заторский Н., Розенфельд Н., Ицковиц С. и др. (июнь 2006 г.). «Колебания и изменчивость в системе р53» . Молекулярная системная биология . 2 : 2006.0033. дои : 10.1038/msb4100068 . ПМК 1681500 . ПМИД 16773083 .
- ^ Проктор CJ, Грей DA (август 2008 г.). «Объяснение колебаний и изменчивости в системе p53-Mdm2» . Системная биология BMC . 2 (75): 75. дои : 10.1186/1752-0509-2-75 . ПМЦ 2553322 . ПМИД 18706112 .
- ^ Чонг К.Х., Самарасингхе С., Куласири Д. (декабрь 2013 г.). «Математическое моделирование базальной динамики р53 и реакции на повреждение ДНК». С-фаКС . 259 (20-й Международный конгресс по математическому моделированию и симуляции): 670–6. дои : 10.1016/j.mbs.2014.10.010 . ПМИД 25433195 .
- ^ Чумаков П.М., Иотсова В.С., Георгиев Г.П. (1982). «[Выделение плазмидного клона, содержащего последовательность мРНК мышиного невирусного Т-антигена]». Доклады Академии наук СССР . 267 (5): 1272–5. ПМИД 6295732 .
- ^ Орен М., Левин А.Дж. (январь 1983 г.). «Молекулярное клонирование кДНК, специфичной для мышиного клеточного опухолевого антигена p53» . Труды Национальной академии наук Соединенных Штатов Америки . 80 (1): 56–9. Бибкод : 1983PNAS...80...56O . дои : 10.1073/pnas.80.1.56 . ПМК 393308 . ПМИД 6296874 .
- ^ Закут-Хури Р., Орен М., Биенц Б. и др. (1983). «Один ген и псевдоген клеточного опухолевого антигена p53». Природа . 306 (5943): 594–7. Бибкод : 1983Natur.306..594Z . дои : 10.1038/306594a0 . ПМИД 6646235 . S2CID 4325094 .
- ^ Закут-Хури Р., Биенц-Тадмор Б., Гивол Д. и др. (май 1985 г.). «Человеческий клеточный опухолевый антиген p53: последовательность и экспрессия кДНК в клетках COS» . Журнал ЭМБО . 4 (5): 1251–5. дои : 10.1002/j.1460-2075.1985.tb03768.x . ПМЦ 554332 . ПМИД 4006916 .
- ^ Бейкер С.Дж., Фирон Э.Р., Нигро Дж.М. и др. (апрель 1989 г.). «Делеции 17 хромосомы и мутации гена p53 при колоректальном раке». Наука . 244 (4901): 217–21. Бибкод : 1989Sci...244..217B . дои : 10.1126/science.2649981 . ПМИД 2649981 .
- ^ Финлей Калифорния, Хиндс П.В., Левин А.Дж. (июнь 1989 г.). «Протоонкоген р53 может действовать как супрессор трансформации» . Клетка . 57 (7): 1083–93. дои : 10.1016/0092-8674(89)90045-7 . ПМИД 2525423 .
- ^ Рэйкрофт Л., Ву ХИ, Лозано Дж. (август 1990 г.). «Активация транскрипции диким типом, но не трансформирующими мутантами антионкогена p53» . Наука . 249 (4972): 1049–1051. Бибкод : 1990Sci...249.1049R . дои : 10.1126/science.2144364 . ПМЦ 2935288 . ПМИД 2144364 .
- ^ Мальцман В., Чижик Л. (сентябрь 1984 г.). «УФ-облучение стимулирует уровни клеточного опухолевого антигена p53 в нетрансформированных клетках мыши» . Молекулярная и клеточная биология . 4 (9): 1689–94. дои : 10.1128/mcb.4.9.1689 . ПМЦ 368974 . ПМИД 6092932 .
- ^ Кастан М.Б., Куэрбитц С.Дж. (декабрь 1993 г.). «Контроль ареста G1 после повреждения ДНК» . Перспективы гигиены окружающей среды . 101 (Приложение 5): 55–8. дои : 10.2307/3431842 . JSTOR 3431842 . ПМЦ 1519427 . ПМИД 8013425 .
- ^ Кошланд Д.Э. (декабрь 1993 г.). «Молекула года». Наука . 262 (5142): 1953. Бибкод : 1993Sci...262.1953K . дои : 10.1126/science.8266084 . ПМИД 8266084 .
- ^ Вено С., Маратрат М., Дюрей С. и др. (август 1998 г.). «Потребность в функциональном домене p53, богатом пролином, для опосредования апоптоза коррелирует со специфической трансактивацией гена PIG3 и с репрессией транскрипции» . Журнал ЭМБО . 17 (16): 4668–79. дои : 10.1093/emboj/17.16.4668 . ПМЦ 1170796 . ПМИД 9707426 .
- ^ Jump up to: а б Ларсен С., Йокочи Т., Исогай Э. и др. (февраль 2010 г.). «LMO3 взаимодействует с p53 и ингибирует его транскрипционную активность». Связь с биохимическими и биофизическими исследованиями . 392 (3): 252–7. дои : 10.1016/j.bbrc.2009.12.010 . ПМИД 19995558 .
- ^ Хармс К.Л., Чен X (март 2005 г.). «С-конец белков семейства р53 является определяющим фактором судьбы клетки» . Молекулярная и клеточная биология . 25 (5): 2014–30. дои : 10.1128/MCB.25.5.2014-2030.2005 . ПМК 549381 . ПМИД 15713654 .
- ^ Белл С., Кляйн С., Мюллер Л. и др. (октябрь 2002 г.). «p53 содержит большие неструктурированные области в своем исходном состоянии». Журнал молекулярной биологии . 322 (5): 917–27. дои : 10.1016/S0022-2836(02)00848-3 . ПМИД 12367518 .
- ^ Цимер М.А., Мейсон А., Карлсон Д.М. (сентябрь 1982 г.). «Бесклеточные трансляции мРНК белков, богатых пролином» . Журнал биологической химии . 257 (18): 11176–80. дои : 10.1016/S0021-9258(18)33948-6 . ПМИД 7107651 .
- ^ Чжу Дж., Чжан С., Цзян Дж. и др. (декабрь 2000 г.). «Определение функциональных доменов р53, необходимых для индукции апоптоза» . Журнал биологической химии . 275 (51): 39927–34. дои : 10.1074/jbc.M005676200 . ПМИД 10982799 .
- ^ Jump up to: а б Хан Дж.М., Пак Б.Дж., Пак С.Г. и др. (август 2008 г.). «AIMP2/p38, каркас для мульти-тРНК-синтетазного комплекса, реагирует на генотоксический стресс через p53» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (32): 11206–11. Бибкод : 2008PNAS..10511206H . дои : 10.1073/pnas.0800297105 . ПМК 2516205 . ПМИД 18695251 .
- ^ Jump up to: а б Койич С., Медеот Е., Гуччионе Е. и др. (май 2004 г.). «Белок Ankrd2, связь между саркомером и ядром скелетных мышц». Журнал молекулярной биологии . 339 (2): 313–25. дои : 10.1016/j.jmb.2004.03.071 . ПМИД 15136035 .
- ^ Jump up to: а б Гувен Н., Бешерель О.Дж., Киджас А.В. и др. (май 2004 г.). «Апратаксин, новый белок, защищающий от генотоксического стресса» . Молекулярная генетика человека . 13 (10): 1081–93. дои : 10.1093/hmg/ddh122 . ПМИД 15044383 .
- ^ Jump up to: а б Фаббро М., Сэвидж К., Хобсон К. и др. (июль 2004 г.). «Комплексы BRCA1-BARD1 необходимы для фосфорилирования p53Ser-15 и остановки G1/S после повреждения ДНК, вызванного ионизирующим излучением» . Журнал биологической химии . 279 (30): 31251–8. дои : 10.1074/jbc.M405372200 . ПМИД 15159397 .
- ^ Jump up to: а б с Ким С.Т., Лим Д.С., Канман С.Э. и др. (декабрь 1999 г.). «Специфичность субстратов и идентификация предполагаемых субстратов членов семейства ATM-киназ» . Журнал биологической химии . 274 (53): 37538–43. дои : 10.1074/jbc.274.53.37538 . ПМИД 10608806 .
- ^ Кан Дж., Фергюсон Д., Сонг Х. и др. (январь 2005 г.). «Функциональное взаимодействие H2AX, NBS1 и p53 в АТМ-зависимых реакциях на повреждение ДНК и подавлении опухоли» . Молекулярная и клеточная биология . 25 (2): 661–70. дои : 10.1128/MCB.25.2.661-670.2005 . ПМК 543410 . ПМИД 15632067 .
- ^ Ханна К.К., Китинг К.Е., Козлов С. и др. (декабрь 1998 г.). «ATM связывается с p53 и фосфорилирует его: картирование области взаимодействия». Природная генетика . 20 (4): 398–400. дои : 10.1038/3882 . ПМИД 9843217 . S2CID 23994762 .
- ^ Вестфаль С.Х., Шмальц С., Роуэн С. и др. (май 1997 г.). «Генетические взаимодействия между atm и p53 влияют на клеточную пролиферацию и контрольные точки клеточного цикла, индуцированные облучением». Исследования рака . 57 (9): 1664–7. ПМИД 9135004 .
- ^ Стельзл Ю., Ворм Ю., Лаловски М. и др. (сентябрь 2005 г.). «Сеть белок-белкового взаимодействия человека: ресурс для аннотирования протеома» . Клетка . 122 (6): 957–68. дои : 10.1016/j.cell.2005.08.029 . hdl : 11858/00-001M-0000-0010-8592-0 . ПМИД 16169070 .
- ^ Ян С., Ван Х., Бойд Д.Д. (март 2002 г.). «ATF3 подавляет экспрессию коллагеназы IV типа (MMP-2) массой 72 кДа, противодействуя p53-зависимой трансактивации промотора коллагеназы» . Журнал биологической химии . 277 (13): 10804–12. дои : 10.1074/jbc.M112069200 . ПМИД 11792711 .
- ^ Чен С.С., Чанг ПК, Ченг Ю.В. и др. (сентябрь 2002 г.). «Подавление онкогенной активности STK15 требует независимой от трансактивации функции р53» . Журнал ЭМБО . 21 (17): 4491–9. дои : 10.1093/emboj/cdf409 . ПМК 126178 . ПМИД 12198151 .
- ^ Леу Дж.И., Дюмон П., Хафи М. и др. (май 2004 г.). «Митохондриальный p53 активирует Bak и вызывает разрушение комплекса Bak-Mcl1». Природная клеточная биология . 6 (5): 443–50. дои : 10.1038/ncb1123 . ПМИД 15077116 . S2CID 43063712 .
- ^ Jump up to: а б с д и ж Донг Ю, Хакими М.А., Чен X и др. (ноябрь 2003 г.). «Регуляция BRCC, голоферментного комплекса, содержащего BRCA1 и BRCA2, с помощью сигналосомоподобной субъединицы и ее роль в репарации ДНК» . Молекулярная клетка . 12 (5): 1087–99. дои : 10.1016/S1097-2765(03)00424-6 . ПМИД 14636569 .
- ^ Jump up to: а б с Сенгупта С., Роблес А.И., Линке С.П. и др. (сентябрь 2004 г.). «Функциональное взаимодействие между хеликазой BLM и 53BP1 в Chk1-опосредованном пути во время ареста S-фазы» . Журнал клеточной биологии . 166 (6): 801–13. дои : 10.1083/jcb.200405128 . ПМК 2172115 . ПМИД 15364958 .
- ^ Ван XW, Ценг А., Эллис Н.А. и др. (август 2001 г.). «Функциональное взаимодействие р53 и ДНК-хеликазы BLM при апоптозе» . Журнал биологической химии . 276 (35): 32948–55. дои : 10.1074/jbc.M103298200 . ПМИД 11399766 .
- ^ Гаркавцев ИВ, Клей Н, Григорян ИА и др. (декабрь 2001 г.). «Белок синдрома Блума взаимодействует и сотрудничает с р53 в регуляции транскрипции и контроле роста клеток». Онкоген . 20 (57): 8276–80. дои : 10.1038/sj.onc.1205120 . ПМИД 11781842 . S2CID 13084911 .
- ^ Jump up to: а б Ян Ц., Чжан Р., Ван XW и др. (август 2002 г.). «Обработка соединений Холлидея хеликазами BLM и WRN регулируется р53» . Журнал биологической химии . 277 (35): 31980–7. дои : 10.1074/jbc.M204111200 . hdl : 10026.1/10341 . ПМИД 12080066 .
- ^ Абрамович С., Вернер Х. (2003). «Функциональные и физические взаимодействия между BRCA1 и p53 в регуляции транскрипции гена IGF-IR». Гормональные и метаболические исследования . 35 (11–12): 758–62. дои : 10.1055/s-2004-814154 . ПМИД 14710355 . S2CID 20898175 .
- ^ Оучи Т., Монтейро А.Н., Август А. и др. (март 1998 г.). «BRCA1 регулирует p53-зависимую экспрессию генов» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (5): 2302–6. Бибкод : 1998PNAS...95.2302O . дои : 10.1073/pnas.95.5.2302 . ЧВК 19327 . ПМИД 9482880 .
- ^ Чай Ю.Л., Цуй Дж., Шао Н. и др. (январь 1999 г.). «Второй домен BRCT белков BRCA1 взаимодействует с p53 и стимулирует транскрипцию с промотора p21WAF1/CIP1». Онкоген . 18 (1): 263–8. дои : 10.1038/sj.onc.1202323 . ПМИД 9926942 . S2CID 7462625 .
- ^ Чжан Х., Сомасундарам К., Пэн Ю. и др. (апрель 1998 г.). «BRCA1 физически связывается с p53 и стимулирует его транскрипционную активность». Онкоген . 16 (13): 1713–21. дои : 10.1038/sj.onc.1201932 . ПМИД 9582019 . S2CID 24616900 .
- ^ Марморштейн Л.Ю., Оучи Т., Ааронсон С.А. (ноябрь 1998 г.). «Продукт гена BRCA2 функционально взаимодействует с p53 и RAD51» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (23): 13869–74. Бибкод : 1998PNAS...9513869M . дои : 10.1073/pnas.95.23.13869 . ПМК 24938 . ПМИД 9811893 .
- ^ Урамото Х., Идзуми Х., Нагатани Г. и др. (апрель 2003 г.). «Физическое взаимодействие опухолевого супрессора p53/p73 с CCAAT-связывающим транскрипционным фактором 2 (CTF2) и дифференциальная регуляция экспрессии гена группы 1 человека с высокой подвижностью (HMG1)» . Биохимический журнал . 371 (Часть 2): 301–10. дои : 10.1042/BJ20021646 . ПМЦ 1223307 . ПМИД 12534345 .
- ^ Jump up to: а б Ли Л., Юнгман М., Диксон Дж.Э. (январь 2000 г.). «Человеческие фосфатазы Cdc14 взаимодействуют с белком-супрессором опухоли p53 и дефосфорилируют его» . Журнал биологической химии . 275 (4): 2410–4. дои : 10.1074/jbc.275.4.2410 . ПМИД 10644693 .
- ^ Лучани М.Г., Хатчинс Дж.Р., Желева Д. и др. (июль 2000 г.). «С-концевой регуляторный домен р53 содержит функциональный сайт стыковки циклина А». Журнал молекулярной биологии . 300 (3): 503–18. дои : 10.1006/jmbi.2000.3830 . ПМИД 10884347 .
- ^ Абабне М., Гетц С., Монтенарх М. (май 2001 г.). «Понижение активности протеинкиназы cdc2/циклин B путем связывания p53 с p34 (cdc2)». Связь с биохимическими и биофизическими исследованиями . 283 (2): 507–12. дои : 10.1006/bbrc.2001.4792 . ПМИД 11327730 .
- ^ Абедини М.Р., Мюллер Э.Дж., Брун Дж. и др. (июнь 2008 г.). «Цисплатин индуцирует p53-зависимое убиквитинирование FLICE-подобного ингибирующего белка в клетках рака яичников» . Исследования рака . 68 (12): 4511–7. дои : 10.1158/0008-5472.CAN-08-0673 . ПМИД 18559494 .
- ^ Jump up to: а б Гудлок Д.М., Цзян К., Перейра Э. и др. (август 2003 г.). «Регуляторные взаимодействия между киназой контрольной точки Chk1 и белками ДНК-зависимого протеинкиназного комплекса» . Журнал биологической химии . 278 (32): 29940–7. дои : 10.1074/jbc.M301765200 . ПМИД 12756247 .
- ^ Тиан Х., Фадже А.Т., Ли С.Л. и др. (2002). «Радиационно-индуцированное фосфорилирование Chk1 по S345 связано с p53-зависимыми путями остановки клеточного цикла» . Неоплазия . 4 (2): 171–80. дои : 10.1038/sj.neo.7900219 . ПМК 1550321 . ПМИД 11896572 .
- ^ Чжао Л., Сэмюэлс Т., Винклер С. и др. (январь 2003 г.). «Циклин G1 обладает ингибирующей рост активностью, связанной с путями супрессора опухолей ARF-Mdm2-p53 и pRb». Молекулярные исследования рака . 1 (3): 195–206. ПМИД 12556559 .
- ^ Jump up to: а б Ито А., Кавагути Ю., Лай Ч. и др. (ноябрь 2002 г.). «Для его деградации необходимо деацетилирование р53, опосредованное MDM2-HDAC1» . Журнал ЭМБО . 21 (22): 6236–45. дои : 10.1093/emboj/cdf616 . ПМК 137207 . ПМИД 12426395 .
- ^ Jump up to: а б Ливенгуд Дж.А., Скоггин К.Е., Ван Орден К. и др. (март 2002 г.). «Транскрипционная активность p53 опосредуется через взаимодействующий с SRC1 домен CBP/p300» . Журнал биологической химии . 277 (11): 9054–61. дои : 10.1074/jbc.M108870200 . ПМИД 11782467 .
- ^ Jump up to: а б Гиблер Х.А., Лемассон И., Нюборг Дж.К. (июль 2000 г.). «Привлечение p53 CREB-связывающего белка, опосредованное фосфорилированным CREB: новый путь регуляции опухолевого супрессора» . Молекулярная и клеточная биология . 20 (13): 4849–58. дои : 10.1128/MCB.20.13.4849-4858.2000 . ПМК 85936 . ПМИД 10848610 .
- ^ Jump up to: а б Шнайдер Э., Монтенар М., Вагнер П. (ноябрь 1998 г.). «Регуляция активности киназы CAK с помощью p53». Онкоген . 17 (21): 2733–41. дои : 10.1038/sj.onc.1202504 . ПМИД 9840937 . S2CID 6281777 .
- ^ Jump up to: а б Ко Л.Дж., Ши С.Ю., Чен X и др. (декабрь 1997 г.). «p53 фосфорилируется CDK7-циклином H p36MAT1-зависимым образом» . Молекулярная и клеточная биология . 17 (12): 7220–9. дои : 10.1128/mcb.17.12.7220 . ПМК 232579 . ПМИД 9372954 .
- ^ Явузер Ю., Смит Г.К., Блисс Т. и др. (июль 1998 г.). «Независимая от конца ДНК активация ДНК-РК, опосредованная ассоциацией с ДНК-связывающим белком C1D» . Гены и развитие . 12 (14): 2188–99. дои : 10.1101/gad.14.12.2188 . ПМК 317006 . ПМИД 9679063 .
- ^ Jump up to: а б Ризос Х., Дифенбах Э., Бадхвар П. и др. (февраль 2003 г.). «Ассоциация p14ARF с репрессором транскрипции p120E4F усиливает ингибирование клеточного цикла» . Журнал биологической химии . 278 (7): 4981–9. дои : 10.1074/jbc.M210978200 . ПМИД 12446718 .
- ^ Сэнди П., Гостисса М., Фогал В. и др. (январь 2000 г.). «p53 участвует в остановке роста, опосредованной p120E4F» . Онкоген . 19 (2): 188–99. дои : 10.1038/sj.onc.1203250 . ПМИД 10644996 .
- ^ Jump up to: а б с Галлахер В.М., Аргентини М., Сьерра В. и др. (июнь 1999 г.). «MBP1: новый мутантный белок-партнер, специфичный для p53, с онкогенными свойствами» . Онкоген . 18 (24): 3608–16. дои : 10.1038/sj.onc.1202937 . ПМИД 10380882 .
- ^ Каддихи А.Р., Вонг А.Х., Тэм Н.В. и др. (апрель 1999 г.). «Двухцепочечная РНК-активированная протеинкиназа PKR физически связывается с белком-супрессором опухоли p53 и фосфорилирует p53 человека по серину 392 in vitro». Онкоген . 18 (17): 2690–702. дои : 10.1038/sj.onc.1202620 . ПМИД 10348343 . S2CID 22467088 .
- ^ Синобу Н., Маэда Т., Асо Т. и др. (июнь 1999 г.). «Физическое взаимодействие и функциональный антагонизм между фактором элонгации РНК-полимеразы II и p53» . Журнал биологической химии . 274 (24): 17003–10. дои : 10.1074/jbc.274.24.17003 . ПМИД 10358050 .
- ^ Гроссман С.Р., Перес М., Кунг А.Л. и др. (октябрь 1998 г.). «Комплексы p300/MDM2 участвуют в MDM2-опосредованной деградации p53» . Молекулярная клетка . 2 (4): 405–15. дои : 10.1016/S1097-2765(00)80140-9 . ПМИД 9809062 .
- ^ Ан В., Ким Дж., Редер Р.Г. (июнь 2004 г.). «Упорядоченные совместные функции PRMT1, p300 и CARM1 при активации транскрипции с помощью p53» . Клетка . 117 (6): 735–48. дои : 10.1016/j.cell.2004.05.009 . ПМИД 15186775 .
- ^ Пасторчич М., Дас Х.К. (ноябрь 2000 г.). «Регуляция транскрипции гена пресенилина-1 человека с помощью факторов транскрипции ets и протоонкогена p53» . Журнал биологической химии . 275 (45): 34938–45. дои : 10.1074/jbc.M005411200 . ПМИД 10942770 .
- ^ Jump up to: а б Ван XW, Йе Х., Шеффер Л. и др. (июнь 1995 г.). «Модуляция p53 активности эксцизионной репарации нуклеотидов, связанной с TFIIH» . Природная генетика . 10 (2): 188–95. дои : 10.1038/ng0695-188 . hdl : 1765/54884 . ПМИД 7663514 . S2CID 38325851 .
- ^ Ю А, Фань Х.И., Ляо Д. и др. (май 2000 г.). «Активация р53 или потеря белка репарации группы B синдрома Коккейна вызывает метафазную хрупкость человеческих генов U1, U2 и 5S» . Молекулярная клетка . 5 (5): 801–10. дои : 10.1016/S1097-2765(00)80320-2 . ПМИД 10882116 .
- ^ Цай Р.Ю., Маккей Р.Д. (декабрь 2002 г.). «Ядрышковый механизм, контролирующий пролиферацию клеток в стволовых и раковых клетках» . Гены и развитие . 16 (23): 2991–3003. дои : 10.1101/gad.55671 . ЧВК 187487 . ПМИД 12464630 .
- ^ Пэн Ю.К., Куо Ф., Брейдинг Д.Э. и др. (сентябрь 2001 г.). «AMF1 (GPS2) модулирует трансактивацию р53» . Молекулярная и клеточная биология . 21 (17): 5913–24. дои : 10.1128/MCB.21.17.5913-5924.2001 . ПМЦ 87310 . ПМИД 11486030 .
- ^ Вочарасит П., Бижур Г.Н., Змиевский Дж.В. и др. (июнь 2002 г.). «Прямое, активирующее взаимодействие между киназой гликогенсинтазы-3бета и р53 после повреждения ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (12): 7951–5. Бибкод : 2002PNAS...99.7951W . дои : 10.1073/pnas.122062299 . ПМК 123001 . ПМИД 12048243 .
- ^ Jump up to: а б Акакура С., Ёсида М., Йонеда Й. и др. (май 2001 г.). «Роль Hsc70 в регуляции нуклеоцитоплазматического транспорта чувствительного к температуре p53 (p53Val-135)» . Журнал биологической химии . 276 (18): 14649–57. дои : 10.1074/jbc.M100200200 . ПМИД 11297531 .
- ^ Ван С., Чен Дж. (январь 2003 г.). «Фосфорилирование и связывание hsp90 опосредуют стабилизацию p53 при тепловом шоке» . Журнал биологической химии . 278 (3): 2066–71. дои : 10.1074/jbc.M206697200 . ПМИД 12427754 .
- ^ Пэн Ю, Чен Л, Ли С и др. (ноябрь 2001 г.). «Ингибирование MDM2 с помощью hsp90 способствует стабилизации мутантного р53» . Журнал биологической химии . 276 (44): 40583–90. дои : 10.1074/jbc.M102817200 . ПМИД 11507088 .
- ^ Чен Д., Ли М., Луо Дж. и др. (апрель 2003 г.). «Прямое взаимодействие между HIF-1 альфа и Mdm2 модулирует функцию p53» . Журнал биологической химии . 278 (16): 13595–8. дои : 10.1074/jbc.C200694200 . ПМИД 12606552 .
- ^ Рави Р., Мукерджи Б., Бхуджвалла З.М. и др. (январь 2000 г.). «Регуляция опухолевого ангиогенеза посредством p53-индуцированной деградации индуцируемого гипоксией фактора 1альфа» . Гены и развитие . 14 (1): 34–44. дои : 10.1101/gad.14.1.34 . ПМК 316350 . ПМИД 10640274 .
- ^ Ханссон Л.О., Фридлер А., Фрейнд С. и др. (август 2002 г.). «Два мотива последовательности HIF-1альфа связываются с сайтом связывания ДНК р53» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (16): 10305–9. Бибкод : 2002PNAS...9910305H . дои : 10.1073/pnas.122347199 . ПМК 124909 . ПМИД 12124396 .
- ^ РГ, Канекал М., Саймон М.К. и др. (март 1998 г.). «Стабилизация p53 дикого типа с помощью индуцируемого гипоксией фактора 1альфа». Природа . 392 (6674): 405–8. Бибкод : 1998Natur.392..405A . дои : 10.1038/32925 . ПМИД 9537326 . S2CID 4423081 .
- ^ Кондо С., Лу Ю., Деббас М. и др. (апрель 2003 г.). «Характеристика клеток и мышей, нацеленных на гены, с дефицитом p53-связывающей киназы, взаимодействующей с гомеодоменом протеинкиназы 1 (HIPK1)» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (9): 5431–6. Бибкод : 2003PNAS..100.5431K . дои : 10.1073/pnas.0530308100 . ПМК 154362 . ПМИД 12702766 .
- ^ Хофманн Т.Г., Мёллер А., Сирма Х. и др. (январь 2002 г.). «Регуляция активности р53 путем его взаимодействия с протеинкиназой-2, взаимодействующей с гомеодоменом». Природная клеточная биология . 4 (1): 1–10. дои : 10.1038/ncb715 . ПМИД 11740489 . S2CID 37789883 .
- ^ Ким Э.Дж., Пак Дж.С., Ум С.Дж. (август 2002 г.). «Идентификация и характеристика HIPK2, взаимодействующего с p73 и модулирующего функции семейства p53 in vivo» . Журнал биологической химии . 277 (35): 32020–8. дои : 10.1074/jbc.M200153200 . ПМИД 11925430 .
- ^ Имамура Т., Изуми Х., Нагатани Г. и др. (март 2001 г.). «Взаимодействие с р53 усиливает связывание модифицированной цисплатином ДНК высокомобильным белком группы 1» . Журнал биологической химии . 276 (10): 7534–40. дои : 10.1074/jbc.M008143200 . ПМИД 11106654 .
- ^ Динтильяк А., Бернюес Дж. (март 2002 г.). «HMGB1 взаимодействует со многими явно неродственными белками, распознавая короткие аминокислотные последовательности» . Журнал биологической химии . 277 (9): 7021–8. дои : 10.1074/jbc.M108417200 . hdl : 10261/112516 . ПМИД 11748221 .
- ^ Вадхва Р., Ягучи Т., Хасан М.К. и др. (апрель 2002 г.). «Член семейства Hsp70, mot-2/mthsp70/GRP75, связывается с доменом цитоплазматической секвестрации белка p53». Экспериментальные исследования клеток . 274 (2): 246–53. дои : 10.1006/excr.2002.5468 . ПМИД 11900485 .
- ^ Стеффан Дж.С., Казанцев А., Спасич-Бошович О. и др. (июнь 2000 г.). «Белок болезни Хантингтона взаимодействует с p53 и CREB-связывающим белком и подавляет транскрипцию» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (12): 6763–8. Бибкод : 2000PNAS...97.6763S . дои : 10.1073/pnas.100110097 . ЧВК 18731 . ПМИД 10823891 .
- ^ Люнг К.М., По Л.С., Цанг Ф.К. и др. (сентябрь 2002 г.). «Кандидатный супрессор опухоли ING1b может стабилизировать р53, нарушая регуляцию р53 с помощью MDM2». Исследования рака . 62 (17): 4890–3. ПМИД 12208736 .
- ^ Гаркавцев И., Григорян И.А., Оссовская В.С. и др. (январь 1998 г.). «Кандидатный супрессор опухоли p33ING1 взаимодействует с p53 в контроле роста клеток». Природа . 391 (6664): 295–8. Бибкод : 1998Natur.391..295G . дои : 10.1038/34675 . ПМИД 9440695 . S2CID 4429461 .
- ^ Jump up to: а б Шисеки М., Нагасима М., Педо Р.М. и др. (май 2003 г.). «p29ING4 и p28ING5 связываются с p53 и p300 и усиливают активность p53». Исследования рака . 63 (10): 2373–8. ПМИД 12750254 .
- ^ Цай К.В., Ценг ХК, Лин В.К. (октябрь 2008 г.). «Два события колебательного сплайсинга влияют на субъядерную локализацию и деградацию белка ING4». Экспериментальные исследования клеток . 314 (17): 3130–41. дои : 10.1016/j.yexcr.2008.08.002 . ПМИД 18775696 .
- ^ Чанг Н.С. (март 2002 г.). «Неанкириновый С-конец икаппы-бальфы физически взаимодействует с р53 in vivo и диссоциирует в ответ на апоптотический стресс, гипоксию, повреждение ДНК и подавление роста, опосредованное трансформирующим фактором роста бета 1» . Журнал биологической химии . 277 (12): 10323–31. дои : 10.1074/jbc.M106607200 . ПМИД 11799106 .
- ^ Jump up to: а б Курки С., Латонен Л., Лайхо М. (октябрь 2003 г.). «Клеточный стресс и повреждение ДНК вызывают различные во времени комплексы Mdm2, p53 и PML и специфическую ядерную релокализацию» . Журнал клеточной науки . 116 (Часть 19): 3917–25. дои : 10.1242/jcs.00714 . ПМИД 12915590 .
- ^ Jump up to: а б Фриман Д.Д., Ли А.Г., Вэй Дж. и др. (февраль 2003 г.). «Супрессор опухоли PTEN регулирует уровни и активность белка p53 посредством фосфатазозависимых и независимых механизмов» . Раковая клетка . 3 (2): 117–30. дои : 10.1016/S1535-6108(03)00021-7 . ПМИД 12620407 .
- ^ Jump up to: а б Чжан Ю, Сюн Ю, Ярбро В.Г. (март 1998 г.). «ARF способствует деградации MDM2 и стабилизирует p53: делеция локуса ARF-INK4a нарушает пути подавления опухоли как Rb, так и p53» . Клетка . 92 (6): 725–34. дои : 10.1016/S0092-8674(00)81401-4 . ПМИД 9529249 .
- ^ Бадчионг Дж.К., Хаас А.Л. (декабрь 2002 г.). «MdmX представляет собой убиквитинлигазу RING-пальца, способную синергически усиливать убиквитинирование Mdm2» . Журнал биологической химии . 277 (51): 49668–75. дои : 10.1074/jbc.M208593200 . ПМИД 12393902 .
- ^ Шварц А., Базуин М., Деккер П. и др. (июль 1997 г.). «Выделение и идентификация человеческого гомолога нового p53-связывающего белка, Mdmx» (PDF) . Геномика . 43 (1): 34–42. дои : 10.1006/geno.1997.4775 . hdl : 2066/142231 . ПМИД 9226370 . S2CID 11794685 .
- ^ Фраде Р., Бальбо М., Барель М. (декабрь 2000 г.). «RB18A, ген которого локализован на хромосоме 17q12-q21.1, регулирует трансактивирующую активность р53 in vivo». Исследования рака . 60 (23): 6585–9. ПМИД 11118038 .
- ^ Дране П., Барель М., Бальбо М. и др. (декабрь 1997 г.). «Идентификация RB18A, нового регуляторного белка р53 массой 205 кДа, который разделяет антигенные и функциональные свойства с р53» . Онкоген . 15 (25): 3013–24. дои : 10.1038/sj.onc.1201492 . ПМИД 9444950 .
- ^ Ху MC, Цю WR, Ван Ю.П. (ноябрь 1997 г.). «JNK1, JNK2 и JNK3 представляют собой N-концевые сериновые 34 киназы p53» . Онкоген . 15 (19): 2277–87. дои : 10.1038/sj.onc.1201401 . ПМИД 9393873 .
- ^ Лин Ю., Хохлачев А., Фигейс Д. и др. (декабрь 2002 г.). «Связанный со смертью белок 4 связывает MST1 и усиливает апоптоз, индуцированный MST1» . Журнал биологической химии . 277 (50): 47991–8001. дои : 10.1074/jbc.M202630200 . ПМИД 12384512 .
- ^ Таниура Х., Мацумото К., Ёсикава К. (июнь 1999 г.). «Физические и функциональные взаимодействия супрессора роста нейронов некдина с p53» . Журнал биологической химии . 274 (23): 16242–8. дои : 10.1074/jbc.274.23.16242 . ПМИД 10347180 .
- ^ Дэниели Ю., Димитрова Д.Д., Боровец Ю.А. (август 2002 г.). «Стресс-зависимая мобилизация нуклеолина, опосредованная образованием комплекса р53-нуклеолин» . Молекулярная и клеточная биология . 22 (16): 6014–22. дои : 10.1128/MCB.22.16.6014-6022.2002 . ПМК 133981 . ПМИД 12138209 .
- ^ Колалука И.Н., Тосони Д., Нусифоро П. и др. (январь 2008 г.). «NUMB контролирует активность опухолевого супрессора p53». Природа . 451 (7174): 76–80. Бибкод : 2008Natur.451...76C . дои : 10.1038/nature06412 . ПМИД 18172499 . S2CID 4431258 .
- ^ Jump up to: а б с Чой М.К., Мовасса М., Сиггенс Л. и др. (июнь 2010 г.). «Высокопроизводительное секвенирование идентифицирует STAT3 как ДНК-ассоциированный фактор экспрессии генов, зависимой от комплекса p53-NF-kappaB, при сердечной недостаточности у человека» . Геномная медицина . 2 (6): 37. дои : 10,1186/гм158 . ПМК 2905097 . ПМИД 20546595 .
- ^ Jump up to: а б Чжан Ю., Вольф Г.В., Бхат К. и др. (декабрь 2003 г.). «Рибосомальный белок L11 отрицательно регулирует онкопротеин MDM2 и опосредует p53-зависимый путь рибосомально-стрессовой контрольной точки» . Молекулярная и клеточная биология . 23 (23): 8902–12. дои : 10.1128/MCB.23.23.8902-8912.2003 . ПМК 262682 . ПМИД 14612427 .
- ^ Николаев А.Ю., Ли М., Пушкаш Н. и др. (январь 2003 г.). «Parc: цитоплазматический якорь р53» . Клетка . 112 (1): 29–40. дои : 10.1016/S0092-8674(02)01255-2 . ПМИД 12526791 .
- ^ Маланга М., Плешке Дж.М., Клечковска Х.Е. и др. (май 1998 г.). «Поли(АДФ-рибоза) связывается со специфическими доменами р53 и изменяет его функции связывания ДНК» . Журнал биологической химии . 273 (19): 11839–43. дои : 10.1074/jbc.273.19.11839 . ПМИД 9565608 .
- ^ Кахьо Т., Нисида Т., Ясуда Х. (сентябрь 2001 г.). «Участие PIAS1 в сумойлировании опухолевого супрессора p53» . Молекулярная клетка . 8 (3): 713–8. дои : 10.1016/S1097-2765(01)00349-5 . ПМИД 11583632 .
- ^ Вульф Г.М., Лиу Ю.К., Рио А. и др. (декабрь 2002 г.). «Роль Pin1 в регуляции стабильности p53 и трансактивации p21, а также контрольных точек клеточного цикла в ответ на повреждение ДНК» . Журнал биологической химии . 277 (50): 47976–9. дои : 10.1074/jbc.C200538200 . ПМИД 12388558 .
- ^ Закки П., Гостисса М., Учида Т. и др. (октябрь 2002 г.). «Пролилизомераза Pin1 раскрывает механизм контроля функций р53 после генотоксических инсультов». Природа . 419 (6909): 853–7. Бибкод : 2002Natur.419..853Z . дои : 10.1038/nature01120 . ПМИД 12397362 . S2CID 4311658 .
- ^ Хуанг С.М., Шёнталь А.Х., Столлкап MR (апрель 2001 г.). «Усиление p53-зависимой активации гена с помощью коактиватора транскрипции Zac1». Онкоген . 20 (17): 2134–43. дои : 10.1038/sj.onc.1204298 . ПМИД 11360197 . S2CID 21331603 .
- ^ Се С., Ву Х, Ван Ц и др. (ноябрь 2001 г.). «Plk3 функционально связывает повреждение ДНК с остановкой клеточного цикла и апоптозом, по крайней мере частично, через путь p53» . Журнал биологической химии . 276 (46): 43305–12. дои : 10.1074/jbc.M106050200 . ПМИД 11551930 .
- ^ Бахасси Э.М., Конн К.В., Майер Д.Л. и др. (сентябрь 2002 г.). «Поло-подобная киназа 3 млекопитающих (Plk3) представляет собой многофункциональный белок, участвующий в путях реакции на стресс». Онкоген . 21 (43): 6633–40. дои : 10.1038/sj.onc.1205850 . ПМИД 12242661 . S2CID 24106070 .
- ^ Саймонс А., Меламед-Бессудо С., Волкович Р. и др. (январь 1997 г.). «ПАКТ: клонирование и характеристика клеточного белка, связывающего р53, который взаимодействует с Rb» . Онкоген . 14 (2): 145–55. дои : 10.1038/sj.onc.1200825 . ПМИД 9010216 .
- ^ Фусаро Г., Дасгупта П., Растоги С. и др. (ноябрь 2003 г.). «Прогибин индуцирует транскрипционную активность р53 и экспортируется из ядра при передаче апоптотического сигнала» . Журнал биологической химии . 278 (48): 47853–61. дои : 10.1074/jbc.M305171200 . ПМИД 14500729 .
- ^ Фогал В., Гостисса М., Сэнди П. и др. (ноябрь 2000 г.). «Регуляция активности р53 в ядерных тельцах с помощью специфической изоформы PML» . Журнал ЭМБО . 19 (22): 6185–95. дои : 10.1093/emboj/19.22.6185 . ПМК 305840 . ПМИД 11080164 .
- ^ Го А., Саломони П., Луо Дж. и др. (октябрь 2000 г.). «Функция PML в p53-зависимом апоптозе». Природная клеточная биология . 2 (10): 730–6. дои : 10.1038/35036365 . ПМИД 11025664 . S2CID 13480833 .
- ^ Jump up to: а б Чжан З, Чжан Р (март 2008 г.). «Протеосомный активатор гамма PA28 регулирует р53, усиливая его деградацию, опосредованную MDM2» . Журнал ЭМБО . 27 (6): 852–64. дои : 10.1038/emboj.2008.25 . ПМК 2265109 . ПМИД 18309296 .
- ^ Лим С.Т., Чен С.Л., Лим Ю. и др. (январь 2008 г.). «Ядерный FAK способствует пролиферации и выживанию клеток за счет деградации p53, усиленной FERM» . Молекулярная клетка . 29 (1): 9–22. doi : 10.1016/j.molcel.2007.11.031 . ПМК 2234035 . ПМИД 18206965 .
- ^ Бернал Дж.А., Луна Р., Эспина А. и др. (октябрь 2002 г.). «Человеческий секурин взаимодействует с р53 и модулирует р53-опосредованную транскрипционную активность и апоптоз». Природная генетика . 32 (2): 306–11. дои : 10.1038/ng997 . ПМИД 12355087 . S2CID 1770399 .
- ^ Штюрцбехер Х.В., Донзельманн Б., Хеннинг В. и др. (апрель 1996 г.). «p53 напрямую связан с процессами гомологичной рекомбинации посредством взаимодействия белков RAD51/RecA» . Журнал ЭМБО . 15 (8): 1992–2002. дои : 10.1002/j.1460-2075.1996.tb00550.x . ПМК 450118 . ПМИД 8617246 .
- ^ Буххоп С., Гибсон М.К., Ван XW и др. (октябрь 1997 г.). «Взаимодействие р53 с белком Rad51 человека» . Исследования нуклеиновых кислот . 25 (19): 3868–74. дои : 10.1093/нар/25.19.3868 . ПМК 146972 . ПМИД 9380510 .
- ^ Ленг Р.П., Лин Ю., Ма В. и др. (март 2003 г.). «Pirh2, p53-индуцированная убиквитин-белковая лигаза, способствует деградации p53» . Клетка . 112 (6): 779–91. дои : 10.1016/S0092-8674(03)00193-4 . ПМИД 12654245 .
- ^ Шэн Ю., Лайстер Р.К., Лемак А. и др. (декабрь 2008 г.). «Молекулярные основы убиквитилирования p53, опосредованного Pirh2» . Структурная и молекулярная биология природы . 15 (12): 1334–42. дои : 10.1038/nsmb.1521 . ПМК 4075976 . ПМИД 19043414 .
- ^ Романова Л.Ю., Виллерс Х., Благосклонный М.В. и др. (декабрь 2004 г.). «Взаимодействие р53 с репликационным белком А опосредует подавление гомологичной рекомбинации». Онкоген . 23 (56): 9025–33. дои : 10.1038/sj.onc.1207982 . ПМИД 15489903 . S2CID 23482723 .
- ^ Рива Ф., Зуко В., Винк А.А. и др. (декабрь 2001 г.). «Индуцированное УФ-излучением разрез ДНК и рекрутирование ядерного антигена пролиферирующих клеток в места репарации происходят независимо от взаимодействия белка А репликации p53 в клетках p53 дикого типа и мутантных клетках карциномы яичника». Канцерогенез . 22 (12): 1971–8. doi : 10.1093/carcin/22.12.1971 . ПМИД 11751427 .
- ^ Линь Дж., Ян К., Ян З. и др. (август 2004 г.). «Ингибирование S100B восстанавливает уровни p53 в клетках первичного злокачественного рака меланомы» . Журнал биологической химии . 279 (32): 34071–7. дои : 10.1074/jbc.M405419200 . ПМИД 15178678 .
- ^ Jump up to: а б Минти А., Дюмон Х., Кагад М. и др. (ноябрь 2000 г.). «Ковалентная модификация p73альфа с помощью SUMO-1. Двухгибридный скрининг с помощью p73 идентифицирует новые белки, взаимодействующие с SUMO-1, и мотив взаимодействия SUMO-1» . Журнал биологической химии . 275 (46): 36316–23. дои : 10.1074/jbc.M004293200 . ПМИД 10961991 .
- ^ Jump up to: а б Иванчук С.М., Мондал С., Рутка Дж.Т. (июнь 2008 г.). «p14ARF взаимодействует с DAXX: влияние на HDM2 и p53» . Клеточный цикл . 7 (12): 1836–50. дои : 10.4161/cc.7.12.6025 . ПМИД 18583933 .
- ^ Jump up to: а б Ли Д., Ким Дж.В., Со Т. и др. (июнь 2002 г.). «Комплекс SWI/SNF взаимодействует с опухолевым супрессором р53 и необходим для активации р53-опосредованной транскрипции» . Журнал биологической химии . 277 (25): 22330–7. дои : 10.1074/jbc.M111987200 . ПМИД 11950834 .
- ^ Янг Пи Джей, Дэй ПМ, Чжоу Дж и др. (январь 2002 г.). «Прямое взаимодействие между белком выживающего мотонейрона и р53 и его связь со спинальной мышечной атрофией» . Журнал биологической химии . 277 (4): 2852–9. дои : 10.1074/jbc.M108769200 . ПМИД 11704667 .
- ^ Сето Э., Ушева А., Замбетти Г.П. и др. (декабрь 1992 г.). «Р53 дикого типа связывается с ТАТА-связывающим белком и подавляет транскрипцию» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (24): 12028–32. Бибкод : 1992PNAS...8912028S . дои : 10.1073/pnas.89.24.12028 . ПМК 50691 . ПМИД 1465435 .
- ^ Цвекл А., Кашанчи Ф., Брейди Дж.Н. и др. (июнь 1999 г.). «Взаимодействие Pax-6 с белком, связывающим ТАТА-бокс, и белком ретинобластомы». Исследовательская офтальмология и визуальные науки . 40 (7): 1343–50. ПМИД 10359315 .
- ^ Макферсон Л.А., Локтев А.В., Вейгель Р.Дж. (ноябрь 2002 г.). «Опухолосупрессорная активность AP2альфа, опосредованная прямым взаимодействием с p53» . Журнал биологической химии . 277 (47): 45028–33. дои : 10.1074/jbc.M208924200 . ПМИД 12226108 .
- ^ Соренсен Т.С., Гирлинг Р., Ли К.В. и др. (октябрь 1996 г.). «Функциональное взаимодействие DP-1 и p53» . Молекулярная и клеточная биология . 16 (10): 5888–95. дои : 10.1128/mcb.16.10.5888 . ПМК 231590 . ПМИД 8816502 .
- ^ Грин Д.Р., Чипук Дж.Е. (июль 2006 г.). «p53 и метаболизм: внутри TIGAR» . Клетка . 126 (1): 30–2. дои : 10.1016/j.cell.2006.06.032 . ПМИД 16839873 .
- ^ Гобер С., Складановски А., Ларсен А.К. (август 1999 г.). «Взаимодействие между р53 и ДНК-топоизомеразой I регулируется по-разному в клетках с диким типом и мутантным р53» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (18): 10355–60. Бибкод : 1999PNAS...9610355G . дои : 10.1073/pnas.96.18.10355 . ЧВК 17892 . ПМИД 10468612 .
- ^ Мао Ю., Мель И.Р., Мюллер М.Т. (февраль 2002 г.). «Субъядерное распределение топоизомеразы I связано с продолжающейся транскрипцией и статусом р53» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1235–40. Бибкод : 2002PNAS...99.1235M . дои : 10.1073/pnas.022631899 . ПМК 122173 . ПМИД 11805286 .
- ^ Jump up to: а б Коуэлл И.Г., Окороков А.Л., Каттс С.А. и др. (февраль 2000 г.). «Человеческие топоизомеразы IIальфа и IIбета взаимодействуют с С-концевой областью р53». Экспериментальные исследования клеток . 255 (1): 86–94. дои : 10.1006/excr.1999.4772 . PMID 10666337 .
- ^ Derbyshire DJ, Basu BP, Serpell LC и др. (июль 2002 г.). «Кристаллическая структура доменов BRCT человека 53BP1, связанных с опухолевым супрессором p53» . Журнал ЭМБО . 21 (14): 3863–72. дои : 10.1093/emboj/cdf383 . ПМК 126127 . ПМИД 12110597 .
- ^ Экблад CM, Фридлер А, Вепринцев Д и др. (март 2004 г.). «Сравнение BRCT-доменов BRCA1 и 53BP1: биофизический анализ» . Белковая наука . 13 (3): 617–25. дои : 10.1110/ps.03461404 . ПМК 2286730 . ПМИД 14978302 .
- ^ Ло К.В., Кан Х.М., Чан Л.Н. и др. (март 2005 г.). «Легкая цепь динеина массой 8 кДа связывается с р53-связывающим белком 1 и опосредует накопление р53 в ядре, вызванное повреждением ДНК» . Журнал биологической химии . 280 (9): 8172–9. дои : 10.1074/jbc.M411408200 . ПМИД 15611139 .
- ^ Джу В.С., Джеффри П.Д., Кантор С.Б. и др. (март 2002 г.). «Структура региона BRCT 53BP1, связанного с p53, и ее сравнение со структурой BRCT Brca1» . Гены и развитие . 16 (5): 583–93. дои : 10.1101/gad.959202 . ПМК 155350 . ПМИД 11877378 .
- ^ Derbyshire DJ, Basu BP, Date T и др. (октябрь 2002 г.). «Очистка, кристаллизация и предварительный рентгеновский анализ доменов BRCT человеческого 53BP1, связанных с опухолевым супрессором p53». Акта Кристаллографика Д. 58 (Часть 10, Часть 2): 1826–9. Бибкод : 2002AcCrD..58.1826D . дои : 10.1107/S0907444902010910 . ПМИД 12351827 .
- ^ Jump up to: а б Ивабучи К., Бартель П.Л., Ли Б. и др. (июнь 1994 г.). «Два клеточных белка, которые связываются с р53 дикого типа, но не с мутантным» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (13): 6098–102. Бибкод : 1994PNAS...91.6098I . дои : 10.1073/pnas.91.13.6098 . ПМЦ 44145 . ПМИД 8016121 .
- ^ Наумовски Л., Клири М.Л. (июль 1996 г.). «Р53-связывающий белок 53BP2 также взаимодействует с Bc12 и препятствует развитию клеточного цикла на уровне G2/M» . Молекулярная и клеточная биология . 16 (7): 3884–92. дои : 10.1128/MCB.16.7.3884 . ПМК 231385 . ПМИД 8668206 .
- ^ Томасини Р., Самир А.А., Кэрриер А и др. (сентябрь 2003 г.). «TP53INP1 и протеинкиназа-2, взаимодействующая с гомеодоменом (HIPK2), являются партнерами в регуляции активности p53» . Журнал биологической химии . 278 (39): 37722–9. дои : 10.1074/jbc.M301979200 . ПМИД 12851404 .
- ^ Окамура С., Аракава Х., Танака Т. и др. (июль 2001 г.). «p53DINP1, ген, индуцируемый p53, регулирует p53-зависимый апоптоз» . Молекулярная клетка . 8 (1): 85–94. дои : 10.1016/S1097-2765(01)00284-2 . ПМИД 11511362 .
- ^ Ли Л., Ляо Дж., Руланд Дж. и др. (февраль 2001 г.). «Регуляторный контур TSG101/MDM2 модулирует деградацию MDM2 и контроль обратной связи MDM2/p53» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (4): 1619–24. Бибкод : 2001PNAS...98.1619L . дои : 10.1073/pnas.98.4.1619 . ПМК 29306 . ПМИД 11172000 .
- ^ Ляхович А, депутат Шехара (апрель 2003 г.). «Образование супрамолекулярного комплекса между Rad6 и белками пути p53 во время ответа, вызванного повреждением ДНК» . Молекулярная и клеточная биология . 23 (7): 2463–75. дои : 10.1128/MCB.23.7.2463-2475.2003 . ПМК 150718 . ПМИД 12640129 .
- ^ Шен З., Пардингтон-Пуртимун П.Е., Комо Дж.К. и др. (октябрь 1996 г.). «Ассоциации UBE2I с белками RAD52, UBL1, p53 и RAD51 в двугибридной системе дрожжей» . Геномика . 37 (2): 183–6. дои : 10.1006/geno.1996.0540 . ПМИД 8921390 .
- ^ Бернье-Вилламор В., Сэмпсон Д.А., Матунис М.Дж. и др. (февраль 2002 г.). «Структурная основа E2-опосредованной конъюгации SUMO, выявленная комплексом между убиквитин-конъюгирующим ферментом Ubc9 и RanGAP1» . Клетка . 108 (3): 345–56. дои : 10.1016/S0092-8674(02)00630-X . ПМИД 11853669 .
- ^ Сехат Б., Андерссон С., Гирнита Л. и др. (июль 2008 г.). «Идентификация c-Cbl как новой лигазы для рецептора инсулиноподобного фактора роста-I с отличной от Mdm2 ролью в убиквитинировании и эндоцитозе рецептора». Исследования рака . 68 (14): 5669–77. дои : 10.1158/0008-5472.CAN-07-6364 . ПМИД 18632619 .
- ^ Сонг М.С., Сонг С.Дж., Ким С.Ю. и др. (июль 2008 г.). «Супрессор опухоли RASSF1A способствует самоубиквитинированию MDM2, разрушая комплекс MDM2-DAXX-HAUSP» . Журнал ЭМБО . 27 (13): 1863–74. дои : 10.1038/emboj.2008.115 . ПМЦ 2486425 . ПМИД 18566590 .
- ^ Ян В., Дикер Д.Т., Чен Дж. и др. (март 2008 г.). «CARPs усиливают оборот p53 за счет разрушения 14-3-3сигмы и стабилизации MDM2» . Клеточный цикл . 7 (5): 670–82. дои : 10.4161/cc.7.5.5701 . ПМИД 18382127 .
- ^ Абэ Ю., Ода-Сато Э., Тобиуме К. и др. (март 2008 г.). «Передача сигналов Hedgehog подавляет подавление опухоли, опосредованное p53, путем активации Mdm2» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (12): 4838–43. Бибкод : 2008PNAS..105.4838A . дои : 10.1073/pnas.0712216105 . ПМК 2290789 . ПМИД 18359851 .
- ^ Домесен С., Кеппель М., Доббельштейн М. (январь 2008 г.). «Специфическое ингибирование неддилирования, опосредованного Mdm2, с помощью Tip60». Клеточный цикл . 7 (2): 222–31. дои : 10.4161/cc.7.2.5185 . ПМИД 18264029 . S2CID 8023403 .
- ^ Ли М., Чен Д., Шайло А. и др. (апрель 2002 г.). «Дубиквитинирование p53 с помощью HAUSP является важным путем стабилизации p53». Природа . 416 (6881): 648–53. Бибкод : 2002Natur.416..648L . дои : 10.1038/nature737 . ПМИД 11923872 . S2CID 4389394 .
- ^ Брош Р.М., Кармакар П., Соммерс Дж.А. и др. (сентябрь 2001 г.). «p53 модулирует экзонуклеазную активность белка синдрома Вернера» . Журнал биологической химии . 276 (37): 35093–102. дои : 10.1074/jbc.M103332200 . ПМИД 11427532 .
- ^ Чанг Н.С., Пратт Н., Хит Дж. и др. (февраль 2001 г.). «Индукция гиалуронидазой оксидоредуктазы, содержащей домен WW, которая усиливает цитотоксичность фактора некроза опухоли» . Журнал биологической химии . 276 (5): 3361–70. дои : 10.1074/jbc.M007140200 . ПМИД 11058590 .
- ^ Окамото Т., Изуми Х., Имамура Т. и др. (декабрь 2000 г.). «Прямое взаимодействие р53 с белком, связывающим Y-бокс, YB-1: механизм регуляции экспрессии генов человека». Онкоген . 19 (54): 6194–202. дои : 10.1038/sj.onc.1204029 . ПМИД 11175333 . S2CID 19222684 .
- ^ Келли К.Д., Миллер К.Р., Тодд А. и др. (май 2010 г.). «YPEL3, ген, регулируемый р53, который вызывает клеточное старение» . Исследования рака . 70 (9): 3566–75. дои : 10.1158/0008-5472.CAN-09-3219 . ПМК 2862112 . ПМИД 20388804 .
- ^ Уотерман М.Дж., Ставриди Э.С., Уотерман Дж.Л. и др. (июнь 1998 г.). «АТМ-зависимая активация р53 включает дефосфорилирование и ассоциацию с белками 14-3-3». Природная генетика . 19 (2): 175–8. дои : 10.1038/542 . ПМИД 9620776 . S2CID 26600934 .
- ^ Лю Дж., Гроган Л., Нау М.М. и др. (апрель 2001 г.). «Физическое взаимодействие между p53 и геном первичного ответа Egr-1». Международный журнал онкологии . 18 (4): 863–70. дои : 10.3892/ijo.18.4.863 . ПМИД 11251186 .
- ^ Бай Л., торговец JL (июль 2001 г.). «ZBP-89 способствует остановке роста за счет стабилизации р53» . Молекулярная и клеточная биология . 21 (14): 4670–83. дои : 10.1128/MCB.21.14.4670-4683.2001 . ПМК 87140 . ПМИД 11416144 .
- ^ Ямакучи М., Ловенштейн CJ (март 2009 г.). «МиР-34, SIRT1 и p53: петля обратной связи» . Клеточный цикл . 8 (5): 712–5. дои : 10.4161/cc.8.5.7753 . ПМИД 19221490 .
- ^ Ван Ю, Чжан Дж, Ли Дж и др. (май 2019 г.). «CircRNA_014511 влияет на радиочувствительность мезенхимальных стволовых клеток костного мозга путем связывания с миР-29b-2-5p» . Боснийский журнал фундаментальных медицинских наук . 19 (2): 155–163. дои : 10.17305/bjbms.2019.3935 . ПМЦ 6535393 . ПМИД 30640591 .
Внешние ссылки
[ редактировать ]- «База знаний p53» . Лейн Групп в Институте молекулярной и клеточной биологии (IMCB), Сингапур. Архивировано из оригинала 3 января 2006 г. Проверено 6 апреля 2008 г.
- Запись GeneReviews/NCBI/NIH/UW о синдроме Ли-Фраумени
- ОПУХОЛЕВОЙ БЕЛОК p53 @ OMIM
- p53 восстановление функции
- стр53 @ Атлас генетики и цитогенетики в онкологии и гематологии
- TP53 Джин @ GeneCards
- p53 Новости предоставлены научной организацией
- Гудсел Д.С. (1 июля 2002 г.). «Супрессор опухоли р53» . Молекула месяца . Банк данных белков RCSB . Проверено 6 апреля 2008 г.
- Сусси Т. «Веб-сайт p53» . Проверено 6 апреля 2008 г.
- Living LFS Некоммерческая организация поддержки пациентов с синдромом Ли-Фраумени.
- The George Pantziarka TP53 Trust Группа поддержки из Великобритании для людей с синдромом Ли-Фраумени или другими расстройствами, связанными с TP53.
- База данных IARC TP53 Somatic Mutations, поддерживаемая в IARC, Лион, Магали Оливье.
- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB для Human P53.
- научная анимация конформационные изменения р53 при связывании с ДНК
галерея PDB |
|---|