Рецептор эстрогена
| Рецептор эстрогена 1 (ER-альфа) | |||
|---|---|---|---|
 Димер лиганд-связывающей области ERα ( рендеринг PDB на основе 3ERD ). | |||
| Идентификаторы | |||
| Символ | ESR1 | ||
| Альт. символы | ER-α, NR3A1 | ||
| Ген NCBI | 2099 | ||
| HGNC | 3467 | ||
| Омим | 133430 | ||
| PDB | 1ERE | ||
| Refseq | NM_000125 | ||
| Uniprot | P03372 | ||
| Другие данные | |||
| Локус | Chr | ||
| |||
| Рецептор эстрогена 2 (ER-бета) | |||
|---|---|---|---|
 Дисмер лиганд-связывающей области ERβ ( рендеринг PDB на основе 1U3S ). | |||
| Идентификаторы | |||
| Символ | ESR2 | ||
| Альт. символы | ER-β, NR3A2 | ||
| Ген NCBI | 2100 | ||
| HGNC | 3468 | ||
| Омим | 601663 | ||
| PDB | 1 квартал | ||
| Refseq | NM_001040275 | ||
| Uniprot | Q92731 | ||
| Другие данные | |||
| Локус | Хр. 14 Q21-Q22 | ||
| |||
Рецепторы эстрогена ( ERS ) представляют собой группу белков , обнаруженных внутри клеток . Это рецепторы , которые активируются гормональным эстрогеном ( 17β -эстрадиол ). [ 1 ] Существуют два класса ER: рецепторы ядерного эстрогена ( ERα и ERβ ), которые являются членами ядерных рецепторов семейства внутриклеточных рецепторов и мембранных рецепторов эстрогена (MERS) ( GPER (GPR30), ER -X и G Q -MER ) , которые в основном являются G-белковыми рецепторами . Эта статья относится к первому (ER).
После активации эстрогеном ER способен транспортироваться в ядро и связываться с ДНК, чтобы регулировать активность различных генов (то есть это ДНК-связывающий фактор транскрипции ). Однако он также имеет дополнительные функции, независимые от связывания ДНК. [ 2 ]
В качестве гормональных рецепторов для половых стероидов ( рецепторы стероидных гормонов ), ERS, рецепторы андрогена (ARS) и прогестероны рецепторы (PRS) важны для полового созревания и беременности .
Протеомика
[ редактировать ]Существуют две разные формы рецептора эстрогена, обычно называемые α и β , каждая из которых кодируется отдельным геном ( ESR1 и ESR2 соответственно). Активированные гормонами рецепторы эстрогена образуют димеры , и, поскольку две формы коэкспрессируются во многих типах клеток, рецепторы могут образовывать гомодимеры ERα (αα) или ERβ (ββ) или гетеродимеров ERαβ (αβ). [ 3 ] Альфа и бета-рецептор эстрогена демонстрируют значительную общую гомологию последовательности, и оба состоят из пяти доменов, обозначенных A/B до F (перечисленные на N- до C-конце; номера аминокислотных последовательностей относятся к человеческому ER). [ Цитация необходима ]
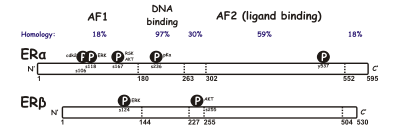
N -концевой домен A/B способен трансактистировать транскрипцию генов в отсутствие связанного лиганда (например, гормон эстрогена). Хотя эта область способна активировать транскрипцию генов без лиганда, эта активация слабая и более селективная по сравнению с активацией, предоставленной доменом E. Домен C, также известный как ДНК-связывающий домен , связывается с элементами ответа эстрогена в ДНК. Домен D представляет собой область шарнира, которая соединяет домены C и E. Домен E содержит полость связывания лиганда, а также сайты связывания для коактиватора и белков корепрессора . Электронный домен в присутствии связанного лиганда способен активировать транскрипцию генов. Функция домена С-концевой F не является полностью ясной и имеет переменную по длине. [ Цитация необходима ]
|
| ||||||||||||||||||||||||||||
Из -за альтернативного сплайсинга РНК известно несколько изоформ ER. По меньшей мере три ERα и пять изоформ ERβ были идентифицированы. Подтипы рецепторов ERβ -изоформ могут трансактистировать транскрипцию только при образовании гетеродимер с функциональным рецептором ERß1 59 кДа. Рецептор ERß3 был обнаружен на высоких уровнях в яичке. Две другие изоформы ERα составляют 36 и 46 кДа. [ 4 ] [ 5 ]
Только у рыб, но не у людей, был описан рецептор ERγ. [ 6 ]
Генетика
[ редактировать ]У людей две формы рецептора эстрогена кодируются различными генами , ESR1 и ESR2 на шестой и четырнадцатой хромосоме (6q25.1 и 14q23.2) соответственно.
Распределение
[ редактировать ]Оба ER широко экспрессируются в разных типах тканей, однако есть некоторые заметные различия в их паттернах экспрессии: [ 7 ]
- ERα , яичниках обнаруживается в эндометрии , клетках рака молочной железы стромальных клеток и гипоталамусе . [ 8 ] У мужчин белок ERα обнаруживается в эпителии эфферентных протоков . [ 9 ]
- Экспрессия белка ERβ была задокументирована в клетках гранулезы яичников , почек , мозга , кости , сердца , [ 10 ] легкие , кишечника слизистая оболочка , простата и эндотелиальные клетки.
Считается, что ERS являются цитоплазматическими рецепторами в их неиспользованном состоянии, но исследования визуализации показали, что только небольшая часть ERS находится в цитоплазме, причем большинство из ER конститутивно в ядре. [ 11 ] Первичная транскрипция «ERα» приводит к нескольким альтернативно сплайсированным вариантам неизвестной функции. [ 12 ]
Лиганды
[ редактировать ]Агонисты
[ редактировать ]- Эндогенные эстрогены (например, эстрадиол , эстрон , эстрал , эстетрол )
- Природные эстрогены (например, конъюгированные эстрогены )
- Синтетические эстрогены (например, этинилэстрадиол , диэтилстилбестрол )
Смешанный (агонист и антагонистический способ действия)
[ редактировать ]- Фитоэстрогены (например, куместрол , даидзеин , генистеин , мироэстрол )
- Селективные модуляторы рецепторов эстрогена (например, тамоксифен , кломифен , ралоксифен )
Антагонисты
[ редактировать ]- Антиэстрогены (например, Fulvestrant , ICI-164384 , Ethamoxytriphetol )
Сродство
[ редактировать ]Связывание и функциональная селективность
[ редактировать ]Домен Helix 12 ER играет решающую роль в определении взаимодействия с коактиваторами и корепрессорами и, следовательно, соответствующего агонистского или антагонистического эффекта лиганда. [ 13 ] [ 14 ]
Различные лиганды могут отличаться по их сродству к альфа и бета -изоформам рецептора эстрогена:
- эстрадиол в равной степени связывается с обоим рецепторами [ 15 ]
- эстрон и ралоксифен связываются преимущественно с альфа -рецептором [ 15 ]
- эстриол и генистеин для бета -рецептора [ 15 ]
подтипа Селективные модуляторы рецепторов эстрогена преимущественно связываются либо с α- или β-мусором рецептора. Кроме того, различные комбинации рецепторов эстрогена могут по -разному реагировать на различные лиганды, что может привести к селективному агонистическим и антагонистическим эффектам тканей. [ 16 ] Было предложено, чтобы соотношение концентрации α- и β-подтипа играет роль при определенных заболеваниях. [ 17 ]
Концепция селективных модуляторов рецепторов эстрогена основана на способности способствовать взаимодействию ER с различными белками, такими как транскрипционный коактиватор или корепрессоры . Кроме того, соотношение коактиватора к белке корепрессора варьируется в разных тканях. [ 18 ] Как следствие, тот же лиганд может быть агонистом в некоторой ткани (где преобладают коактиваторы), в то время как антагонистические в других тканях (где доминируют корепрессоры). Например, тамоксифен является антагонистом в молочной железе и, следовательно, используется в качестве рака молочной железы лечения [ 19 ] но агонист ER в кости (тем самым предотвращающий остеопороз ) и частичный агонист в эндометрии (увеличение риска рака матки ).
Сигнальная трансдукция
[ редактировать ]Поскольку эстроген является стероидным гормоном , он может проходить через фосфолипидные мембраны клетки, и поэтому рецепторы не должны быть связаны с мембраной, чтобы связываться с эстрогеном. [ Цитация необходима ]
Геномный
[ редактировать ]В отсутствие гормона рецепторы эстрогена в основном расположены в цитозоле. Связывание гормонов с рецептором запускает ряд событий, начиная с миграции рецептора из цитозоля в ядро, димеризацию рецептора и последующее связывание димера рецептора с специфическими последовательностями ДНК, известных как элементы отклика гормона . Затем комплекс ДНК/рецепторов рекрутирует другие белки, которые ответственны за транскрипцию нижней ДНК в мРНК и, наконец, белок, что приводит к изменению функции клеток. Рецепторы эстрогена также встречаются в клеточном ядре , и оба подтипа рецептора эстрогена имеют ДНК -связывающий домен и могут функционировать как факторы транскрипции для регулирования продукции белков . [ Цитация необходима ]
Рецептор также взаимодействует с белком активатора 1 и SP-1 для стимулирования транскрипции через несколько коактиваторов, таких как PELP-1 . [ 2 ]
Супрессор опухоли киназы LKB1 коактирует ERα в клеточном ядре посредством прямого связывания, в котором он рекрутирован к промотору ERα-чувствительных генов. Каталитическая активность LKB1 усиливает трансактивацию ERα по сравнению с каталитическим дефицитом LKB1. [ 20 ]
Прямое ацетилирование альфа рецептора эстрогена в остатках лизина в петле с помощью p300 регулирует трансактивацию и чувствительность к гормонам. [ 21 ]
Не геномный
[ редактировать ]Некоторые рецепторы эстрогена ассоциируются с мембраной клеточной поверхности и могут быстро активироваться при воздействии клеток на эстроген. [ 22 ] [ 23 ]
Кроме того, некоторые ER могут ассоциироваться с клеточными мембранами путем прикрепления к кавеолину-1 и формировать комплексы с G-белками , стриатином , рецепторными тирозинкиназами (например, EGFR и IGF-1 ) и нерецепторными тирозинкиназами (EG, SRC ). [ 2 ] [ 22 ] Через стриатину некоторые из этих мембранных, которые могут привести к повышению уровня CA 2+ и оксид азота (нет). [ 24 ] Благодаря рецепторным тирозинкиназам сигналы отправляются в ядро через митоген-активированный протеинкиназой (MAPK/ ERK) путь и путем фосфоинозитид-3-киназы (PL3K/ AKT ). [ 25 ] Гликогенсинтаза-киназа-3 ингибируя фосфорилирование серина (GSK) -3β ингибирует транскрипцию ядерной ER , 118 ядерного ERα. Фосфорилирование GSK-3β удаляет его ингибирующий эффект, и это может быть достигнуто путем PI3K/AKT и пути MAPK/ERK через RSK . [ Цитация необходима ]
Было показано, что 17β-эстрадиол активирует связанный с G GPR30, GPR30 , связанным с G. [ 26 ] Однако субклеточная локализация и роль этого рецептора по -прежнему являются объектом противоречий. [ 27 ]
Различия и неисправность
[ редактировать ]

Рак
[ редактировать ]Рецепторы эстрогена чрезмерно экспрессируются примерно в 70% случаев рака молочной железы , называемых « ER-позитивными », и могут быть продемонстрированы в таких тканях с использованием иммуногистохимии . Были предложены две гипотезы, чтобы объяснить, почему это вызывает онкогенез , и имеющиеся данные свидетельствуют о том, что оба механизма вносят вклад:
- Во -первых, связывание эстрогена с ER стимулирует пролиферацию клеток молочной железы с результирующим увеличением деления клеток и репликации ДНК , что приводит к мутациям.
- Во -вторых, метаболизм эстрогена продуцирует генотоксические отходы.
Результатом обоих процессов является нарушение клеточного цикла , апоптоза и репарации ДНК , что увеличивает вероятность образования опухоли. ERα, безусловно, связан с более дифференцированными опухолями, в то время как доказательства того, что ERβ вовлечен, являются противоречивыми. различные версии гена ESR1 Были идентифицированы (с однонуклеотидными полиморфизмами ) и связаны с различным риском развития рака молочной железы. [ 19 ]
Эстроген и ERS также участвовали в раке молочной железы , раке яичников , раке толстой кишки , раке простаты и раке эндометрия . Усовершенствованный рак толстой кишки связан с потерей ERβ, преобладающим ER в тканях толстой кишки, а рак толстой кишки лечится эРβ-специфическими агонистами. [ 28 ]
Эндокринная терапия рака молочной железы включает в себя селективные модуляторы рецепторов эстрогена (SERM), такие как тамоксифен , которые ведут себя как антагонисты ER в ткани молочной железы или ингибиторы ароматазы , такие как анастрозол . Статус ER используется для определения чувствительности поражений рака молочной железы к ингибиторам тамоксифена и ароматазы. [ 29 ] Другой пост, Raloxifene , использовался в качестве профилактической химиотерапии для женщин, которые, по мнению, имеют высокий риск развития рака молочной железы. [ 30 ] Другой химиотерапевтический антиэстроген, ICI 182 780 (Faslodex), который действует как полный антагонист, также способствует деградации рецептора эстрогена.
Тем не менее, устойчивость de novo к эндокринной терапии подрывает эффективность использования конкурентных ингибиторов, таких как тамоксифен. Гормональная депривация посредством использования ингибиторов ароматазы также станет бесполезной. [ 31 ] Массово параллельное секвенирование генома выявило общее присутствие точечных мутаций на ESR1 , которые являются драйверами для сопротивления, и способствуют агонистской конформации ERα без связанного лиганда . Такая конститутивная, независимая от эстрогена активность обусловлена специфическими мутациями, такими как мутации D538G или Y537S/C/N, в лиганд-связывающем домене ESR1 и способствует пролиферации клеток и прогрессированию опухоли без стимуляции гормонов. [ 32 ]
Менопауза
[ редактировать ]Метаболические эффекты эстрогена у женщин в постменопаузе были связаны с генетическим полиморфизмом бета-бета-рецептора эстрогена (ER-β) . [ 33 ]
Старение
[ редактировать ]Исследования у самок мышей показали, что рецептор-рецептор-альфа-рецептор-альфа-альфа в преоптическом гипоталамусе , когда они стареют. Самки-мыши, которые получали калорийную диету в течение большей части их жизни, поддерживали более высокие уровни ERα в доптическом гипоталамусе, чем их некалорически ограниченные аналоги. [ 8 ]
Ожирение
[ редактировать ]Драматическая демонстрация важности эстрогенов в регуляции осаждения жира происходит от трансгенных мышей , которые были генетически спроектированы с отсутствием функционального гена ароматазы . Эти мыши имеют очень низкий уровень эстрогена и страдают ожирением. [ 34 ] Ожирение также наблюдалось у самок мышей с дефицитом эстрогена, в которых отсутствует рецептор гормонов, стимулирующие фолликулы . [ 35 ] Влияние низкого эстрогена на увеличение ожирения было связано с альфа -рецептором эстрогена. [ 36 ]
Сервики для других целей лечения
[ редактировать ]Середи также изучаются для лечения миомы матки [ 37 ] [ нуждается в обновлении ] и эндометриоз . [ 38 ]
Синдром нечувствительности эстрогена
[ редактировать ]Синдром нечувствительности эстрогена является редким межсексуальным состоянием с 5 сообщенными случаями, в которых рецепторы эстрогена не функционируют. Фенотип . приводит к маскулинизации обширной В отличие от синдрома нечувствительности андрогена , EIS не приводит к изменению пола фенотипа . Это невероятно редко и анологично по отношению к ИИ и формам надпочечников . Причина, по которой AIS распространен, и EIS исключительно редка, заключается в том, что XX AIS не приводит к бесплодию и, следовательно, может быть в материнской линии , в то время как EIS всегда приводит к бесплодию независимо от кариотипа . Отрицательная петля обратной связи между эндокринной системой также встречается в EIS, в котором гонады производят заметно более высокие уровни эстрогена для людей с EIS (119–272 пг/мл xy и 750–3 500 пг/мл XX, см. Средние уровни ), однако нет. Феминизирующие эффекты происходят. [ 39 ] [ 40 ]
Открытие
[ редактировать ]Рецепторы эстрогена были впервые идентифицированы Элвудом В. Дженсеном в Чикагском университете в 1958 году, [ 41 ] [ 42 ] за которого Дженсен был удостоен премии Ласкер . [ 43 ] Ген для второго рецептора эстрогена (ERβ) был идентифицирован в 1996 году Kuiper et al. В простате и яичнике крысы с использованием вырожденных праймеров Eralpha. [ 44 ]
Смотрите также
[ редактировать ]- Мембранный рецептор эстрогена
- Синдром нечувствительности эстрогена
- Дефицит ароматазы
- Ароматазный избыточный синдром
Ссылки
[ редактировать ]- ^ Dahlman-Wright K, Cavailles V, Fuqua SA, Jordan VC, Katzenellenbogen JA, Korach KS, et al. (Декабрь 2006 г.). «Международный союз фармакологии. Lxiv. Рецепторы эстрогена». Фармакологические обзоры . 58 (4): 773–81. doi : 10.1124/pr.58.4.8 . PMID 17132854 . S2CID 45996586 .
- ^ Jump up to: а беременный в Левин ER (август 2005 г.). «Интеграция экстрануклеарных и ядерных действий эстрогена» . Молекулярная эндокринология . 19 (8): 1951–9. doi : 10.1210/me.2004-0390 . PMC 1249516 . PMID 15705661 .
- ^ Li X, Huang J, Yi P, Bambara RA, Hilf R, Muyan M (сентябрь 2004 г.). «Одноцепочечные рецепторы эстрогена (ERS) показывают, что гетеродимер Eralpha/Beta эмулирует функции димера Eralpha в сигнальных путях геномного эстрогена» . Молекулярная и клеточная биология . 24 (17): 7681–94. doi : 10.1128/mcb.24.17.7681-7694.2004 . PMC 506997 . PMID 15314175 .
- ^ Nilsson S, Mäkelä S, Treuter E, Tujague M, Thomsen J, Andersson G, et al. (Октябрь 2001). «Механизмы действия эстрогена». Физиологические обзоры . 81 (4): 1535–65. doi : 10.1152/physrev.2001.81.4.1535 . PMID 11581496 . S2CID 10223568 .
- ^ Leung YK, Mak P, Hassan S, Ho SM (август 2006 г.). «Изоформы рецептора эстрогена (ER) -бета: ключ к пониманию передачи сигналов ER-бета» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (35): 13162–7. Bibcode : 2006pnas..10313162L . doi : 10.1073/pnas.0605676103 . PMC 1552044 . PMID 16938840 .
- ^ Hawkins MB, Thornton JW, Crews D, Skipper JK, Dotte A, Thomas P (сентябрь 2000). «Идентификация третьего отдельного рецептора эстрогена и реклассификация рецепторов эстрогена в телеостах» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (20): 10751–6. Bibcode : 2000pnas ... 9710751H . doi : 10.1073/pnas.97.20.10751 . PMC 27095 . PMID 11005855 .
- ^ Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS (ноябрь 1997). «Распределение тканей и количественный анализ рецептор-рецептор-альфа-альфа (ERALPHA) и рецептор-бета-бета (ERBETA) рибонуклеиновой кислоты в мыши дикого типа и нокаутированной эральфа» . Эндокринология . 138 (11): 4613–21. doi : 10.1210/en.138.11.4613 . PMID 9348186 .
- ^ Jump up to: а беременный Yaghmaie F, Saeed O, Garan SA, Freitag W, Timiras PS, Sternberg H (Jun 2005). «Ограничение калорий снижает потерю клеток и поддерживает иммунореактивность рецептора эстрогена-альфа в преоптическом гипоталамусе самки мышей B6D2F1» (PDF) . Нейро эндокринологические письма . 26 (3): 197–203. PMID 15990721 .
- ^ Hess RA (июль 2003 г.). «Эстроген в репродуктивном тракте взрослых мужчин: обзор» . Репродуктивная биология и эндокринология . 1 (52): 52. doi : 10.1186/1477-7827-1-52 . PMC 179885 . PMID 12904263 .
- ^ Babiker FA, Windt LJ, Eickels M, Grohe C, Meyer R, Doeennndans PA (февраль 2002 г.). «Астрогенные гормоны Действие в твердости: регуляторная сеть и функция» . Сердечно -сосудистые исследования . 53 (3): 709–19. doi : 10,1016/s0008-6363 (01) 00526-0 . PMID 11861041 .
- ^ Htun H, Holth LT, Walker D, Davie Jr, Hager GL (февраль 1999). «Прямая визуализация рецептора эстрогена человека выявляет роль лиганда в ядерном распределении рецептора» . Молекулярная биология клетки . 10 (2): 471–86. doi : 10.1091/mbc.10.2.471 . PMC 25181 . PMID 9950689 .
- ^ Pfeffer U, Fecarotta E, Vidali G (май 1995). «Коэкспрессия множественных вариантов рецепторов эстрогена мессенджера в нормальных и опухолевых тканях молочной железы и в клетках MCF-7». РАНКА . 55 (10): 2158–65. PMID 7743517 .
- ^ Ascenzi P, Bocedi A, Marino M (август 2006 г.). «Структурно-функциональная связь рецептора эстрогена альфа и бета: воздействие на здоровье человека». Молекулярные аспекты медицины . 27 (4): 299–402. doi : 10.1016/j.mam.2006.07.001 . PMID 16914190 .
- ^ Bourguet W, Germain P, Gronemeyer H (октябрь 2000). «Лиганд-связывающие домены ядерного рецептора: трехмерные структуры, молекулярные взаимодействия и фармакологические последствия». Тенденции в фармакологических науках . 21 (10): 381–8. doi : 10.1016/s0165-6147 (00) 01548-0 . PMID 11050318 .
- ^ Jump up to: а беременный в Чжу Б.Т., Хан Гз, Шим Джи, Вэнь Й., Цзян XR (сентябрь 2006 г.). «Количественная структура-активность взаимосвязи различных эндогенных метаболитов эстрогена для альфа-рецептора человека и бета-подтипов человека: понимание структурных детерминантов, способствующих дифференциальному связыванию подтипа». Эндокринология . 147 (9): 4132–50. doi : 10.1210/en.2006-0113 . PMID 16728493 .
- ^ Кансра С., Ямагата С., Сниде Л., Фостер Л., Бен-Джонатан Н. (июль 2005 г.). «Дифференциальное влияние антагонистов рецепторов эстрогена на пролиферацию лактотрофа и высвобождение пролактина и высвобождение пролактина». Молекулярная и клеточная эндокринология . 239 (1–2): 27–36. doi : 10.1016/j.mce.2005.04.008 . PMID 15950373 . S2CID 42052008 .
- ^ Bakas P, Liapis A, Vlahopoulos S, Giner M, Logotheti S, Creatsas G, et al. (Ноябрь 2008). «Рецептор эстрогена альфа и бета в миоме матки: основа для измененной чувствительности эстрогена» . Плодородие и бесплодия . 90 (5): 1878–85. doi : 10.1016/j.fertnstert.2007.09.019 . HDL : 10442/7330 . PMID 18166184 .
- ^ Shang Y, Brown M (март 2002 г.). «Молекулярные детерминанты для тканевой специфичности сывороток». Наука . 295 (5564): 2465–8. Bibcode : 2002sci ... 295.2465S . doi : 10.1126/science.1068537 . PMID 11923541 . S2CID 30634073 .
- ^ Jump up to: а беременный Deroo BJ, Korach KS (март 2006 г.). «Рецепторы эстрогена и болезнь человека» . Журнал клинических исследований . 116 (3): 561–70. doi : 10.1172/jci27987 . PMC 2373424 . PMID 16511588 .
- ^ Nath-Sain S, Marignani PA (2009-04-15). «Каталитическая активность LKB1 способствует передаче сигналов альфа -рецептора эстрогена» . Молекулярная биология клетки . 20 (11): 2785–2795. doi : 10.1091/mbc.e08-11-1138 . PMC 2688557 . PMID 19369417 .
- ^ Wang C, Fu M, Angeletti RH, Siconolfi-Baez L, Reutens AT, Albanese C, et al. (Май 2001). «Прямое ацетилирование рецепторной петли рецептора эстрогена с помощью p300 регулирует трансактивацию и чувствительность к гормонам» . Журнал биологической химии . 276 (21): 18375–83. doi : 10.1074/jbc.m100800200 . PMID 11279135 .
- ^ Jump up to: а беременный Zivadinovic D, Gametchu B, Watson CS (2005). «Уровни мембранного рецептора эстрогена-альфа в клетках рака молочной железы MCF-7 предсказывают реакции цАМФ и пролиферации» . Исследование рака молочной железы . 7 (1): R101–12. doi : 10.1186/bcr958 . PMC 1064104 . PMID 15642158 .
- ^ Björnström L, Sjöberg M (Jun 2004). «Эстроген-рецептор-зависимая активация AP-1 посредством негеномной передачи сигналов» . Ядерный рецептор . 2 (1): 3. DOI : 10.1186/1478-1336-2-3 . PMC 434532 . PMID 15196329 .
- ^ Lu Q, Pallas DC, Surks HK, Baur WE, Mendelsohn ME, Karas RH (Dec 2004). «Стриатин собирает мембранный сигнальный комплекс, необходимый для быстрой, негеномной активации эндотелиальной NO синтазы с помощью рецептора эстрогена альфа» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (49): 17126–31. Bibcode : 2004pnas..10117126L . doi : 10.1073/pnas.0407492101 . PMC 534607 . PMID 15569929 .
- ^ S, Endoh H, Massoako Y, Week T, Uchiya S, Sasaki H, et al. (Декабрь 1995). Активность. Наука 270 (5241): 1491–4 Bibcode : 1995sci . doi : 10.1126/ Science PMID 7491495 . 4662264S2CID
- ^ Prossnitz ER, Arterburn JB, Sklar LA (февраль 2007 г.). «GPR30: AG-белковой рецептор для эстрогена» . Молекулярная и клеточная эндокринология . 265–266: 138–42. doi : 10.1016/j.mce.2006.12.010 . PMC 1847610 . PMID 17222505 .
- ^ Otto C, Rohde-Schulz B, Schwarz G, Fuchs I, Klewer M, Brittain D, et al. (Октябрь 2008). «G-белок-связанный рецептор 30 локализуется в эндоплазматическом ретикулуме и не активируется эстрадиолом» . Эндокринология . 149 (10): 4846–56. doi : 10.1210/en.2008-0269 . PMID 18566127 .
- ^ Харрис Х.А., Альберт Л.М., Литюрби Ю., Маламас М.С., Мьюшоу Р.Е., Миллер С.П. и др. (Октябрь 2003 г.). «Оценка агониста рецептора эстрогена-бета у животных моделей заболеваний человека» . Эндокринология . 144 (10): 4241–9. doi : 10.1210/en.2003-0550 . PMID 14500559 .
- ^ Clemons M, Danson S, Howell A (август 2002 г.). «Тамоксифен (" nolvadex "): обзор". Обзоры лечения рака . 28 (4): 165–80. doi : 10.1016/s0305-7372 (02) 00036-1 . PMID 12363457 .
- ^ Фабиан К.Дж., Кимлер Б.Ф. (март 2005 г.). «Селективные модуляторы рецептора эстрогена для первичной профилактики рака молочной железы» . Журнал клинической онкологии . 23 (8): 1644–55. doi : 10.1200/jco.2005.11.005 . PMID 15755972 .
- ^ Oesterreich S, Davidson NE (Dec 2013). «Поиск мутаций ESR1 при раке молочной железы» . Природа генетика . 45 (12): 1415–6. doi : 10.1038/ng.2831 . PMC 4934882 . PMID 24270445 .
- ^ Li S, Shen D, Shao J, Crowder R, Liu W, Prat A, et al. (Сентябрь 2013). «Устойчивые к эндокрино-терапии варианты ESR1, выявленные геномной характеристикой ксенотрансплантатов, полученных из молочной железы» . Сотовые отчеты . 4 (6): 1116–30. doi : 10.1016/j.celrep.2013.08.022 . PMC 3881975 . PMID 24055055 .
- ^ Дараби М., Ани М., Панджепур М., Раббани М., Мовахедиан А., Зариан Е. (2011). «Влияние полиморфизма рецептора эстрогена β A1730G на реакцию экспрессии гена ABCA1 на заместительную терапию гормонами в постменопаузе». Генетическое тестирование и молекулярные биомаркеры . 15 (1–2): 11–5. doi : 10.1089/gtmb.2010.0106 . PMID 21117950 .
- ^ Hewitt KN, Boon WC, Murata Y, Jones Me, Simpson ER (сентябрь 2003 г.). «Мышь ароматаза нокаутирует с сексуально диморфным нарушением гомеостаза холестерина» . Эндокринология . 144 (9): 3895–903. doi : 10.1210/en.2003-0244 . PMID 12933663 .
- ^ Данилович Н., Бабу П.С., Син В., Гердес М., Кришнамурти Х, Сайрам М.Р. (ноябрь 2000 г.). «Дефицит эстрогена, ожирение и аномалии скелета у самок мышей, стимулирующих фолликул-стимулирующие гормоны (forko)» . Эндокринология . 141 (11): 4295–308. doi : 10.1210/endo.141.11.7765 . PMID 11089565 .
- ^ Ohlsson C, Hellberg N, Parini P, Vidal O, Bohlooly-Y M, Bohlooly M, et al. (Ноябрь 2000). «Ожирение и нарушенный профиль липопротеинов у мужчин-дефицитных рецепторов эстрогена-альфа-дефицита». Биохимическая и биофизическая исследовательская коммуникация . 278 (3): 640–5. doi : 10.1006/bbrc.2000.3827 . PMID 11095962 .
- ^ Lingxia X, Taixiang W, Xiaoyan C (апрель 2007 г.). Xie L (ред.). «Селективные модуляторы рецепторов эстрогена (SERM) для лейомиомы матки». Кокрановская база данных систематических обзоров (2): CD005287. doi : 10.1002/14651858.cd005287.pub2 . PMID 17443581 .
- ^ Ван Хозель М.Х., Чен Й.Л., Чжэн А., Ван К., Мурад С.М. и др. (Кокрановская гинекология и группа фертильности) (май 2021 г.). «Селективные модуляторы рецепторов эстрогена (SERM) для эндометриоза» . Кокрановская база данных систематических обзоров . 2021 (5): CD011169. doi : 10.1002/14651858.cd011169.pub2 . PMC 8130989 . PMID 33973648 .
- ^ Томас Л. Лемке, Дэвид А. Уильямс (24 января 2012 г.). Принципы Фойи по лекарственной химии . Липпинкотт Уильямс и Уилкинс. С. 1392–. ISBN 978-1-60913-345-0 .
- ^ Смит Эп, Бойд Дж., Фрэнк Г.Р., Такахаши Х., Коэн Р.М., Спекер Б. и др. (Октябрь 1994). «Устойчивость к эстрогенам, вызванная мутацией в гене эстрогена-рецептор у человека» . Новая Англия Журнал медицины . 331 (16): 1056–61. doi : 10.1056/nejm199410203311604 . PMID 8090165 .
- ^ Дженсен Э.В., Джордан ВК (июнь 2003 г.). «Рецептор эстрогена: модель для молекулярной медицины» (аннотация) . Клиническое исследование рака . 9 (6): 1980–9. PMID 12796359 .
- ^ Дженсен Е. (2011). «Разговор с Элвудом Дженсеном. Интервью Дэвида Д. Мур» . Ежегодный обзор физиологии . 74 : 1–11. doi : 10.1146/annurev-physiol-020911-153327 . PMID 21888507 .
- ^ Дэвид Брейси, 2004 г. « Ученый UC выигрывает« Американскую Нобелевскую »исследовательскую премию ». Пресс -релиз Университета Цинциннати.
- ^ Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA (Jun 1996). «Клонирование нового рецептора, экспрессируемого в простате и яичнике крысы» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (12): 5925–30. doi : 10.1073/pnas.93.12.5925 . PMC 39164 . PMID 8650195 .
Внешние ссылки
[ редактировать ]- Рецепторы эстрогена в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Дэвид С. Гудселл (2003-09-01). «Рецептор эстрогена» . Банк данных белков , исследовательский сотрудничество по структурной биоинформатике (RCSB). Архивировано из оригинала 11 марта 2006 года . Получено 2008-03-15 .